Hoofdstuk 3: DE GEMIDDELDE EVOLUTIE
Ga naar : Beginpagina Index van het boek Samenvatting
HOOFDSTUK 3 : DE GEMIDDELDE EVOLUTIE
De gemiddelde evolutie : een heuristisch model
1.1. Verspreiding in de evolutie
1.2. Beperking van de verspreiding
2. INFORMATIE ALS BRON VAN COMPLEXITEIT EN COHERENTIE IN DE EVOLUTIE
2.1. Toevallige selectie in de evolutie
2.2. Informatie en selectie in de evolutie : de invloed van de rode vlag
2.3. Geïnformeerde variaties en geïnformeerde selecties in de evolutie
2.4. Wat is informatie binnen de evolutiecontext ?
3. FLUXMAXIMALISATIE EN ATTRACTOREN IN DE GEMIDDELDE EVOLUTIE
3.1. Demping van variaties en het ontstaan van informatie en attractoren in de gemiddelde evolutie
3.2. Hiërarchische niveau’s in de gemiddelde evolutie
3.3. Fluxmaximalisatie en quasi-evenwicht : de link tussen microscopisch en macroscopisch gedrag
4. FUNDAMENTELE ONZEKERHEID IN DE GEMIDDELDE EVOLUTIE
DE GEMIDDELDE EVOLUTIE :
VAN BIT TOT ATOOM EN ECOSYSTEEM
De gemiddelde evolutie : een heuristisch model
In het vorige hoofdstuk is duidelijk gemaakt dat in de evolutie van ons heelal heel veel toevalligheden een bepalende rol hebben gespeeld. Zelfs in die mate dat het onwaarschijnlijk is dat bijvoorbeeld de huidige fauna en flora zich opnieuw zou ontwikkelen indien de evolutie zich zou herhalen. Al het classificatiewerk van een bioloog als Linnaeus zou daardoor niet relevant zijn. Als Carl Linnaeus nog leefde, zou deze gedachte wellicht een ware nachtmerrie zijn.Als het toeval een dergelijke belangrijke rol speelt in de evolutie, en we willen toch een inzicht krijgen in de basisprincipes van het evolutieproces, moeten we de rol van het toeval trachten te reduceren tot zijn relevante proporties. We zullen dit doen door het uitvoeren van een reusachtig gedachtenexperiment. In plaats van de vraag te stellen of de evolutie zich op dezelfde wijze zou kunnen herhalen, zullen we ervan uitgaan dat dit niet het geval (door de toevallige variaties). We zullen daarom een andere vraag stellen :
"Welke evolutiepatronen zouden we zien als we de evolutie een miljoen maal zouden herhalen ?"
Welke kenmerken van het evolutieproces zouden dan keer op keer terugkomen ?
Deze vraag is de basisvraag waar dit boek een antwoord op tracht te formuleren.
Op het einde van de negentiende eeuw constateerde de Amerikaanse wetenschapper en filosoof Charles Sanders Peirce (1839-1914) dat de "natuur de tendens heeft om gewoontes te vormen". Onder deze gewoontes bevinden zich niet enkel de natuurwetten die de ontwikkeling van de cosmos kunnen verklaren. Het ontstaan van de natuurwetten op zich moet immers kunnen verklaard worden, en ook de natuurwetten zijn langzaam ge-evolueerd.Het is duidelijk dat onze zoektocht naar de "gewoontevorming" in de evolutie van het heelal sterk zal beïnvloed zijn door de context waarin we leven als mens op aarde en door de kennis die we tot op heden hebben verzameld. Niettegenstaande dit feit, is de aard van deze fundamentele vraag zodanig dat ze tot kritisch nadenken stemt over de werkelijk fundamentele aspecten van de evolutie. Alhoewel het antwoord beïnvloed zal zijn door de tijdsgeest, zullen we verplicht zijn om tot de kern van de zaak door te denken. Het resultaat van dit denkproces heb ik
"de gemiddelde evolutie"
genoemd.
In dit hoofdstuk zullen we die aspecten van het evolutieproces beschrijven, waarvan we verwachten dat ze wellicht steeds zouden terugkomen indien we de evolutie konden herhalen. Als gevolg hiervan, zal dit hoofdstuk vrij algemeen en soms wat abstract aandoen.
In de volgende hoofdstukken zal het basismodel van dit hoofdstuk verder verduidelijkt worden voor meer specifieke gevallen. Bij het lezen van de volgende hoofdstukken kan het dan ook soms nuttig zijn om naar dit hoofdstuk terug te grijpen. In de volgende hoofdstukken zal ook meer ingegaan worden op de grondleggers van bepaalde theorieën die hier worden gebruikt.
De nadruk bij het schrijven van dit hoofdstuk en van dit boek ligt op het geven van een integrale visie op het evolutieproces. Vandaar dat aan sommige gangbare begrippen een aangepaste interpretatie wordt gegeven, die toelaat de samenhang van het evolutiemodel te verhogen. Waar nodig, zal het onderscheid tussen de gangbare en de in dit boek gebruikte interpretatie worden verduidelijkt.
Het model dat hier wordt voorgesteld is een "heuristisch model". Het maakt gebruik van bestaande puzzelstukken, om ze te combineren tot een vernieuwde versie van de puzzel. Niet zozeer de inhoud is vernieuwend, doch wel de compositie. Ik noem het een heuristisch model, omdat het open staat voor verdere ontwikkeling en evolutie. Het is het resultaat van een persoonlijke visie en is zeker niet te beschouwen als een pasklare en finale oplossing. Ik nodig de lezer dan ook uit mee te denken, gedurende het doornemen van dit werk.Terug naar de index van dit hoofdstuk
1.1. Verspreiding in de evolutie"Alles stroomt" (Panta rei in het Grieks)
Dit is een basisgedachte van de Griekse wijsgeer Heraclitus. Dit is ook de basis van het ganse evolutieproces.
Het enige constante en universele aan de evolutie is dat het een ononderbroken stroming van materie, energie en informatie is. Het universum kan niet worden gevat door het beschrijven van deelelementen ervan ; er kunnen immers continu nieuwe kenmerken ontstaan of ontdekt worden. Het universum is een onverdeeld en ononderbroken geheel. Elke beschrijving van een deel ervan, doet eigenlijk afbreuk aan de eigenheid van het totale.
Dit gezegd zijnde, zou ik eigenlijk consequent moeten zijn en stoppen met verdere beschrijvingen.
Ik zal echter toch doorgaan, omdat ik ervan uitga dat deze "evolutiestroom" toch beter kan begrepen worden, door het geven van een aantal complementaire beschrijvingen.
We zullen echter een eerste basisgedachte formuleren, die het "panta rei" principe meer specifiek weergeeft.
Kerngedachte
De evolutie is gekenmerkt door een continue kwantitatieve en kwalitatieve verspreiding van materie, energie en informatie.
Het woord stroming is vervangen door "verspreiding". Dit geeft immers weer dat de evolutie niet in 1 welbepaalde richting gaat, maar in alle mogelijke richtingen verandert.
Bovendien is inhoud van de stroming verder gespecifieerd : het is een stroming van materie, energie en informatie. De verspreiding heeft bovendien betrekking op de verspreiding in de ruimte (kwantitatief), als op een verspreiding van de verschijningsvormen (kwalitatief) van deze drie stromingskenmerken.
Hierbij enkele voorbeelden om de gedachten te vestigen :
- verspreiding van energie ("E")
De oerknal is hiervan wellicht het duidelijkste voorbeeld. In een fractie van een seconde is een enorme hoeveelheid energie vrijgekomen. Sindsdien is deze energie continu verspreid over het uitdijende heelal. Sindsdien zijn ook de verschillende energievormen onstaan : de diversiteit aan golflengtes van de stralingen is bijvoorbeeld continu toegenomen.
- verspreiding van materie ("M")
- Als een kind een uur gespeeld heeft in een kinderkamer, is het duidelijk dat er een natuurlijke tendens bestaat tot verspreiding van het speelgoed (de materie) over de kamer (een voorbeeld van een "ruimtelijke verspreiding").
- Ook de diversiteit aan levende organismen is een zichtbaar voorbeeld van zowel de kwalitatieve als kwantitatieve verspreiding van materie.
- Het is duidelijk dat sinds de oerknal alle materie zich verspreid heeft over het heelal. Denk maar aan de ontelbare sterren, sterrenstelsels, enz...
- Een ander duidelijk voorbeeld van verspreiding doet zich voor wanneer we een druppel inkt in een glas water toevoegen : de inkt zal zich geleidelijk aan verspreiden over het water in het glas. Dit gaat door tot het water een homogeen blauwe kleur heeft.
- verspreiding van informatie ("I")
De laatste 2000 jaar is de mens zeer succesvol geweest op het vlak van de verspreiding van informatie over de aarde. Niet alleen is de kennis enorm toegenomen, ook het aantal informatiedragers is sterk gestegen (vb. papier, boeken, CD’s,...) en ook de snelheid van informatieverspreiding is razendsnel toegenomen.
Voor het verdere verloop van het boek zullen we soms afkortingen gebruiken : "I" voor informatie, "M" voor materie en "E" voor energie. Deze afkortingen slaan op alle verschijningsvormen van informatie, materie en energie.
Bovendien weten we dat materie kan omgezet worden in een equivalente hoeveelheid energie, en dat ook het omgekeerde mogelijk is. Hout kunnen we bijvoorbeeld verbranden tot warmte-energie : omgekeerd is bijvoorbeeld veel van de oerknalenergie omgezet in allerhande kerndeeltjes, die de basis vormen voor alle materie die we rondom ons zien. (De uitwisselbaarheid van materie en energie werd door Einstein weergegeven in de formule e=mc2.)
Om reden van die uitwisselbaarheid zullen energie en materie dikwijls samen aangeduid worden met de afkorting "M/E".
Omdat we weten dat de verspreiding van energie, materie en informatie een voortdurende stroom is, kunnen we de eerste kerngedachte als volgt symbolisch voorstellen :
M/Ep à M/Er + M/Ev
Ip à Ir + Iv
Dit betekent dat vanaf de oerknal, alle M/E en I het potentieel had (vandaar het suffix "p") om zich te verspreiden (suffix "v"). Deze verspreiding gebeurde echter niet in één ogenblik, doch gaat continu door. Hierbij worden allerlei verschijningsvormen van M/E en I gerealiseerd (vandaar de "r" van realiseren). Een houtblok vertegenwoordigt bijvoorbeeld een bepaalde hoeveelheid M/Ep : tijdens verbranding wordt een deel M/Ev verspreid (vb. onder de vorm van warmte) en wordt een deel M/Er omgezet in houtskool. Pas als het hout volledig is omgezet in warmte is de M/E-verspreiding volledig ten einde.
1.2. Beperking van de verspreiding
Wanneer we de omgeving rondomons bekijken, zien we tal van organisatievormen die sterk geordend zijn, en die ons weinig doen denken aan de continue toename van de verspreiding.
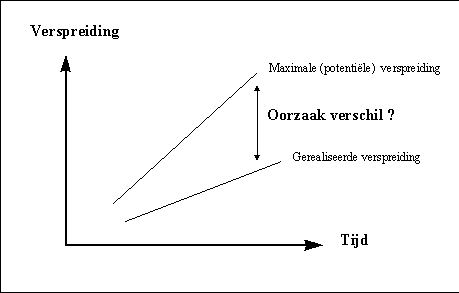
Bij de expansie van het universum, neemt de maximale potentiële verspreiding steeds toe. De werkelijke (gerealiseerde) verspreiding neemt ook steeds toe, doch met een kleinere snelheid. Zoals de onderstaande figuur aantoont, neemt het verschil tussen de werkelijke en de maximale verspreiding ook steeds toe. Het verschil tussen beide is de orde die we waarnemen, bijvoorbeeld onder de vorm van tastbare materie. Met andere woorden, naarmate de verspreiding in het heelal toeneemt, neemt ook de orde toe.
H3F1 : Beperking van de verspreiding (naar Layzer, 1984)
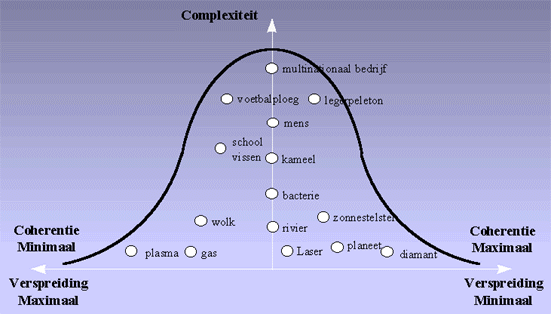
Een cruciale vraag die we zullen moeten stellen is wat de oorzaak is van deze orde. Hoe komt het dat de verspreiding gebeurt via tussenstadia van complexere organisatievormen, in plaats van onmiddellijk tot het maximale niveau van verspreiding te komen ?Dat er een scala van min of meerdere complexere organisatievormen mogelijk zijn, is iets wat we in onze thuisbasis, het zonnestelsel, maar al te duidelijk kunnen waarnemen. We kunnen zeer complexe levende organisatievormen waarnemen, zoals bijvoorbeeld een internationaal bedrijf of een leger en éénvoudigere levensvormen zoals een bacterie.
H3F2 : De complexiteit en coherentie van de verschillende materiële verschijningsvormen verschilt zeer sterk
De niet levende vormen zijn veel minder complex, doch ze verschillen ook zeer sterk in de mate van verspreiding. Een gas is een typevoorbeeld van een materiële vorm die sterk verspreid is. De coherentie tussen de gasmolecules is heel laag. In een diamant daarentegen zijn de atomen heel sterk geordend (zeer coherent), wat de diamant heel sterk maakt, en wat ook zijn hoge mate van symmetrie verklaart. Nochtans is de complexiteit van een diamant veel lager dan die van een bacterie of een school vissen.
Wanneer we dus op zoek gaan naar de oorzaak van de beperking van de verspreiding, moeten we op zoek gaan naar de factoren die aanleiding geven tot de verschillen in coherentie en complexiteit van de verschillende verschijningsvormen rondom ons.Terug naar de index van dit hoofdstuk
2. INFORMATIE ALS BRON VAN COMPLEXITEIT EN COHERENTIE IN DE EVOLUTIE
Informatie krijgt een steeds belangrijker rol toebedeeld in de studie van de evolutie, zowel binnen de fysica als binnen de biologie.
De fysicus John Wheeler stelt dit heel extreem voor :" Het meest elementaire in de beschrijving van de fysica, is het het elementaire quantumfenomeen, waarbij een elementaire ja-nee vraag wordt ontwikkeld. Met andere woorden, elke fysische eenheid heeft zijn bestaan te danken aan bits, of elementaire ja-nee indicaties." Dit is bekend geraakt onder de vorm van zijn gevleugelde uitspraak "it from bit", waarmee hij wilde samenvatten dat alles gebaseerd is op informatie.
Ook in de biologie is de aandacht voor informatie sterk gestegen, zodanig dat er zelfs een aparte wetenschappelijke tak is ontstaan die de "bio-semiotiek" wordt genoemd, of de studie van de (ontwikking van) betekenis van signalen. De bioloog Jesper Hoffmeyer spreekt zelfs van de "semiotisering van de natuur", waarbij de natuur meer en meer wordt aanzien als een geheel dat gekenmerkt wordt door een uitwisseling van signalen (informatie). Vandaar dat ik aan de gevleugelde uitstpraak van Wheeler nog de stelling "fit from bit" zou willen toevoegen, waarmee ik bedoel dat ook de "survival of the fittest" in de biologie een proces is waar de plant of het dier dat de beste historische informatie weet te verzamelen en te gebruiken, ook de grootste kans heeft op overleving.
Dit boek verschilt van vele andere werken over evolutie door de nadruk die het legt op de invloed van informatie, vanwaar de titel "De gemiddelde evolutie : van bit tot atoom en ecosysteem". In vele klassieke werken wordt informatie voorgesteld als iets wat pas heel laat in de evolutie is ontstaan, nadat atomen en ecosystemen zijn ontstaan. Hier wordt een gewijzigde interpratie van het begrip informatie ingevoerd, waaruit volgt dat atomen en ecosystemen het gevolg zijn van het ontstaan van informatie...Als informatie zo belangrijk is voor de gemiddelde evolutie, is het nuttig eens dieper in te gaan op het ontstaan van informatie, en de invloed ervan op de evolutie.
- Een gedachtenexperiment ...
Om de rol van informatie in de evolutie te verduidelijken, zullen we eerst een reeks denkbeeldige experimenten uitvoeren.
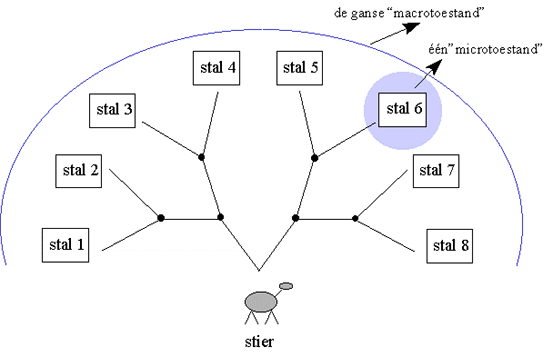
Beeld u zich in dat we over een stier beschikken, die voor een reeks tweesprongen staat. Afhankelijk van de richting die hij kiest bij elke tweesprong, zal hij in één van de verschillende stallen terechtkomen. De stier is zodanig gekozen dat hij geen enkele voorkeur heeft voor de linker of rechter zijde van de tweesprong, doch hij wordt wel onweerstaanbaar aangetrokken wordt door een rode vlag.
Figuur H3F3 : de stier staat voor 3 opeenvolgende tweesprongen : na het nemen van 3 beslissingen komt hij in 1 van de 8 stallen terecht
We starten dus onze reeks gedachtenexperimenten door de stier voor 3 opeenvolgende tweesprongen te plaatsen, zoals aangegeven in het schema hierboven. De stier ziet de volgende tweesprong pas, als hij zijn keuze gemaakt heeft in de vorige.Experiment 1 : geen rode vlag
- We zetten de stier op het startpunt en laten 3 opeenvolgende keuzes maken.
Na het maken van zijn 3 opeenvolgende keuzes, zal hij zal in 1 van de acht stallen terechtkomen.
Welke stal het zal zijn, is niet te voorspellen.Het enige dat we kunnen voorspellen is dat er 1 kans op 8 is dat hij in bijvoorbeeld stal 8 terechtkomt (of in één van de andere stallen).
- Stel dat we 8 stieren zouden hebben en we laten ze één voor één kiezen, zullen ze wellicht tamelijk goed verspreid zijn over de 8 stallen.
- Als we het experiment met de 8 stieren vele malen zouden herhalen, zouden de stieren, gemiddeld gezien, perfect verspreid zijn over de 8 stallen.
Experiment 2 : zeer klein rood vlagje
We herhalen het experiment 1, doch met een lichte wijziging. Op alle knooppunten in de richting van stal 5 plaatsen we een zeer klein rood vlagje van 1 vierkante milimeter (onzichtbaar voor de stier).
Aangezien de stier het vlagje niet ziet (het is te klein), zal hij opnieuw in één van de acht stallen terechtkomen. Opnieuw is niet voorspelbaar welke stal. Ook als we op gelijkaardige wijze experiment b en c herhalen, is de uitkomst hetzelfde als in 1a.
Experiment 3 : klein rood vlagje
- We zetten de stier op het startpunt en laten 3 opeenvolgende keuzes maken.
Op alle knooppunten in de richting van stal 5 plaatsen we een klein rood vlagje van 10
vierkante centimeter . In welke stal de stier nu zal terechtkomen, is niet met zekerheid te zeggen.
Het hangt af of de stier het vlagje telkens ziet. Zoja, zal hij in stal 5 terechtkomen, zoniet kan hij opnieuw in de andere stallen terechtkomen. Stel dat de kans dat hij het vlagje ziet tamelijk groot is (bijvoorbeeld ongeveer 75%) is, is er meer kans dat de stier in stal 5 terechtkomt dan in stal 4 of 6 : er is ook meer kans dat hij in stal 4 terechtkomt dan in stal 3.- Stel dat we 8 stieren zouden hebben en we laten ze één voor één kiezen, zullen ze wellicht niet meer zo goed verspreid zijn over de acht stallen. Er is nu meer kans om meerdere stieren aan te treffen in stal 5.
- Als we het experiment met de 8 stieren vele malen zouden herhalen, zouden de stieren, gemiddeld gezien, met een bepaalde waarschijnlijkheid rond stal 5 geconcentreerd zijn. De verspreiding zal dan perfect (met wiskundige zekerheid) af te leiden zijn uit de waarschijnlijkheid dat de stier de vlag ziet.
Experiment 4 : grote rode vlag
- We zetten de stier op het startpunt en laten 3 opeenvolgende keuzes maken.
Op alle knooppunten in de richting van stal 5 plaatsen we een grote rode vlag van 1 vierkante meter. In welke stal de stier nu zal terechtkomen, is nu wel met zekerheid te zeggen.
Aangezien de stier de rode vlag altijd ziet, en hij er sterk door aangetrokken wordt, zal hij zich telkens naar stal 5 begeven.- Stel dat we 8 stieren zouden hebben en we laten ze één voor één kiezen, zullen ze allemaal in stal 5 terechtkomen. Een slechtere verspreiding (of betere coherentie) is niet mogelijk.
- Als we het experiment met de 8 stieren vele malen zouden herhalen, zouden alle stieren steeds in stal 5 terechtkomen.
Experiment 5 :
a) We voeren experiment 1 uit (zonder vlag) met een stier die al heel veel het experiment 4 heeft uitgevoerd. Zodanig veel, dat hij uit gewoonte opnieuw naar stal 5 loopt, ook al is de vlag niet meer aanwezig. Hij zal dus met zekerheid in stal 5 terechtkomen.
- Stel dat we 8 stieren zouden hebben, getraind als die ene stier, en we laten ze één voor één kiezen, zullen ze allemaal in stal 5 terechtkomen. Een slechtere verspreiding is niet mogelijk.
- Als we het experiment met de 8 stieren vele malen zouden herhalen, zouden alle stieren steeds in stal 5 terechtkomen.
Wanneer we de verspreiding van een groot aantal stieren in experiment 1, 3 en 4 grafisch zouden uitzetten, krijgen we waarschijnlijk onderstaande patronen.
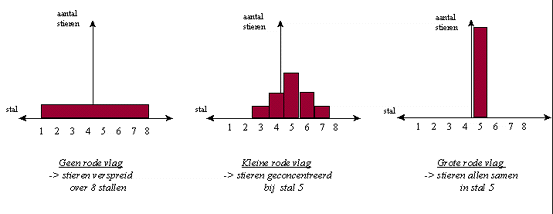
Fig H3F4 : de invloed van de rode vlag in stal 5 op de verspreiding van de stierenHet is duidelijk dat de experimenten met de stier metaforisch zijn voor de gemiddelde evolutie. De gemiddelde evolutie kan immers beschouwd worden als een zich vertakkend netwerk van evolutiewegen. De situatie met de stier toont een voorbeeld van een dergelijk "vertakkend netwerk" met diverse evolutiewegen. De evolutiewegen leiden in dit geval naar de stallen. Het is duidelijk dat de evolutie in het universum tot vele andere resultaten kan leiden.
In de volgende paragrafen zullen we de metafoor naar de gemiddelde evolutie vertalen, en zien wat we uit deze gedachtenexperimenten kunnen leren over verspreiding, informatie, coherentie en complexiteit.Terug naar de index van dit hoofdstuk
2.1. Toevallige selectie in de evolutie
In afwezigheid van de rode vlag (experiment 1) gebeurt de keuze van de richting van de stier bij elke tweesprong puur toevallig. Daardoor verspreiden de stieren zich gelijkmatig over de verschillende stallen. Verspreiding was inderdaad het eerste kenmerk van de gemiddelde evolutie. Bij elke beslissing moet de stier de linkerkant of de rechterkant kiezen. De selectie van één van beide kanten, is onlosmakelijk verbonden met de verspreiding.
De selectie van één van de twee symmetrische zijden, heeft nog een aantal fundamentele eigenschappen, die kenmerkend zijn voor de ganse evolutie.
- Selectie en symmetriebreking :
Vooraleer de stier zijn keuze heeft gemaakt, bevond hij zich in een symmetrische situatie. Na de keuze van één van de twee zijden was deze symmetrie niet meer aanwezig was na de keuze : "de symmetrie wordt verbroken" door elke keuze. Doordat de stier onwetend is over welke keuze hij zal maken, en de keuze toevallig is, kunnen we ook niet voorspellen in welke stal hij zal terechtkomen.
De symmetriebreking kan hierbij aanzien worden als een maat voor de verspreiding. Aanvankelijk was de symmetrie hoog, doch door de verspreiding, is er bij elk beslissingspunt een symmetriebreking gebeurd.
Dit kan ook waargenomen worden wanneer we de evolutie van het heelal beschouwen vanaf de oerknal. Op het moment van de oerknal was de verspreiding van materie, energie en informatie nihil : alles was gebundeld in 1 geheel. Sinds de oerknal heeft een reeks opeenvolgende symmetriebrekingen voor toenemende diversiteit van materie, energie en informatie gezorgd. Deze gebroken symmetrie is zichtbaar in alle verschijningsvormen die we kunnen waarnemen.
- Selectie en reductie/toename van potentieel
Voordat de stier zijn eerste keuze had gemaakt, waren er nog veel potentiele keuzes. Nadat echter de eerste selectie was gebeurd, is een nieuwe reële situatie (nieuwe M/Er, nieuwe M/Ep) ontstaan. Het potentieel aan keuzes, met andere woorden aan mogelijke evoluties en evolutiewegen, is gereduceerd. Merk op dat in de werkelijke evolutie - in tegenstelling tot bij het gedachtenexperiment - , het "aantal stallen" in de loop van de evolutie steeds zal toenemen, waardoor ook de maximaal mogelijke verspreiding toeneemt (meer en meer stieren, zullen zich over meer en meer stallen kunnen verspreiden).- Selectie, symmetrie, herhaling en waarschijnlijkheid
Als we het eerste experiment vele malen herhalen (cfr. 1c), zal de uitkomst echter nog symmetrisch zijn, namelijk de stieren zijn uiteindelijk homogeen (dus symmetrisch) verspreid over de acht stallen. Noemen we 1 stal een "microscopische situatie" of "microtoestand" en het totaalbeeld - de 8 stallen samen - (als bij een kikvorsperspectief) de "macroscopische situatie" of "macrotoestand", dan zien we dat de toevallige verspreiding wel de microscopische symmetrie verbreekt, doch niet volledig de macroscopische.
Herhaling laat toe "de wet van de grote getallen" te gebruiken : het laat ons toe de "gemiddelde evolutie" en de "gemiddelde symmetrie" te voorspellen. Als de toevallige variaties ons beletten om met zekerheid een voorspelling te doen over de keuzes van de stieren, kunnen we door voldoende aantal herhalingen toch een "statistische" of "probabilistische" voorspelling doen. De evolutie is toevallig omdat we onwetend zijn over het verloop ervan : er is niet voldoende informatie om de richting ervan te voorspellen. Dat de evolutie in de richting van een totale verspreiding verloopt, is dus een uiting van twee kenmerken van de evolutie :
- het is een uiting van de rol van het toeval
- is is een uiting van de onwetendheid (ontbreken van informatie) binnen het systeem (in het voorbeeld is de stier onwetend), er is een tekort aan informatie om de evolutie richting te geven.
Belangrijk is dat, bij elke evolutie waar een dergelijk informatietekort is, het systeem zal evolueren naar zowel de maximale verspreiding over de microtoestanden, als de maximale verspreiding over de macrotoestanden. De fundamentele reden is het informatietekort. Er is niets in het systeem dat de stier naar een bepaalde stal aantrekt, of van een bepaalde stal afstoot : het gevolg is dat hij -bij vele herhalingen - zich gemiddeld zal verspreiden over de stallen.
Een duidelijk voorbeeld hiervan vinden we ook bij een de druppel inkt die we in een glas water brengen. Er is geen richtinggevende factor in het water dat de inktmolecules aantrekt of afstoot om geconcentreerd te blijven in hun oorspronkelijke druppelvorm. Daardoor gaan de inktmolecules hun vrije gang en ze verspreiden zich "microscopisch". Het gevolg is dat ze na verloop van tijd volledig verspreid zullen over het ganse glas : met andere woorden ze verspreiden zich ook "macroscopisch".
- Verspreiding is een gevolg van toevallige selectie van evolutierichtingen.
- Bij elk van deze toevallige selecties wordt de symmetrie van het systeem gebroken.
- Bij toevallige selecties is de meest waarschijnlijke toestand, de toestand van de totale verspreiding. Deze meest waarschijnlijke toestand is de toestand waarin het systeem het meest zal waargenomen bij het herhalen van de evolutie, met andere woorden het is ook de stabielste toestand : we noemen dit de toestand van "stabiel evenwicht".
Terug naar de index van dit hoofdstuk
2.2. Informatie en selectie in de evolutie :
de invloed van de rode vlag1) De invloed van de rode vlag :
In afwezigheid van de rode vlag, is de verspreiding perfect. De beslissingen van de stier zijn puur toevallig. De waarschijnlijkheid om in 1 van de acht stallen terecht te komen is even groot. "De selectie is toevallig".
De rode vlag heeft voor de stier een betekenis : hij wordt erdoor aangetrokken. De betekenis van de vlag is "Kom langs hier". De rode vlag is dus informatie voor de stier.
We zien dat de informatie, als ze duidelijk genoeg is, de stier in een bepaalde richting beweegt. Hierdoor is de verspreiding niet meer gelijk over de stallen. Ook bij vele herhalingen is de uitkomst, in tegenstelling tot experiment 1c, niet meer symmetrisch. Met andere woorden, de aanwezigheid van informatie verbreekt, vanaf een kritische waarde (als de informatie duidelijk genoeg is) zowel de microscopische als de macroscopische symmetrie. De informatie beinvloedt de selectie : daarom kan gesproken worden van een "geïnformeerde selectie", in tegenstelling tot de zuiver "toevallige selectie".
De reële situatie heeft bovendien ook de symmetrie van de potentiële situatie gebroken.
2) Leren (experiment 5)
Uit experiment 5 zien we dat informatie kan worden samengevat tot een model of schema van de werkelijkheid. Dit model kan dan opgeslagen worden in een geheugen, in dit geval in het brein van de stier. De stier heeft dus een schema van het traject in zich opgeslagen. Het schema in het brein van de stier is een vereenvoudigde weergave van het traject, waarmee hij de weg naar stal 5 terugvindt.
Informatie opgeslagen in een schema kan de selecterende evolutie beinvloeden.
De aanwezigheid van informatie verhoogt de selectie-waarschijnlijkheid van bepaalde richtingen in de evolutie.
- Naarmate de hoeveelheid informatie kwalitatief en kwantitatief stijgt, neemt de verspreiding die kenmerkend is voor de evolutie binnen een bepaalde context af (met andere woorden, de coherentie neemt toe). De hoeveelheid informatie kan hierbij varieren van bijna nul (bijna totale onwetendheid en totale onzekerheid of dus zonder voorkennis) tot bijna 100% (vrijwel alwetendheid en 100% zekerheid of complete voorkennis). De verspreiding in de evolutie varieert hierbij van nul tot volledig.
- Aangezien de informatie over het totale universum nooit 100% kan zijn, kan informatie enkel binnen een bepaalde context de verspreiding afremmen. Als gevolg hiervan is er in de gemiddelde evolutie een "informatietekort" dat steeds toeneemt, waardoor ook de gemiddelde verspreiding continu blijft toenemen. De gemiddelde evolutie is een verspreidende evolutie.
Terug naar de index van dit hoofdstuk
2.3. Geïnformeerde variaties en geïnformeerde selecties in de evolutie
Nu het duidelijk is geworden dat informatie een belangrijke rol speelt in de evolutie, kunnen we dieper inzoomen op de manier waarop dit gebeurt.
In essentie is de evolutie een opeenvolging van variaties, waarbij nieuwe evolutiewegen worden gevormd, en selecties, waarbij bepaalde evolutiewegen worden geselecteerd.
Het totaal aantal mogelijke variaties komt overeen met de maximaal mogelijke verspreiding.Nu is het zo dat zowel de variaties, als de selecties, als beide kunnen beïnvloed worden door informatie.
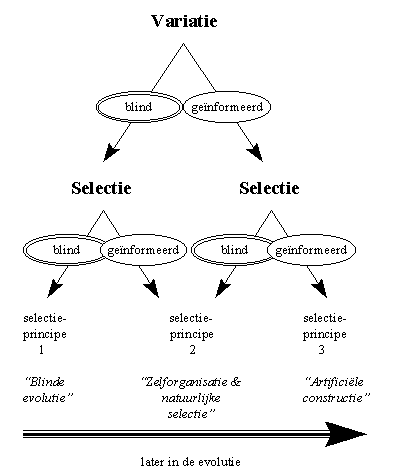
H3F5 : Verschillende selectieprincipes in de natuur
Indien er geen informatie aanwezig is in het systeem dat we beschouwen, krijgen we een volledig "blinde" evolutie, waarbij zowel de variatie als de selectie toevallig gebeuren. Het resultaat komt overeen met de maximale verspreiding, zoals we dit bijvoorbeeld in de atomen van een gas kunnen waarnemen.
In het proces dat Darwin de "natuurlijke selectie" heeft genoemd, wordt uitgegaan van toevallige variaties van de genetische informatie van planten of dieren. Bepaalde van deze variaties zullen leiden tot mutanten die beter geschikt zijn voor de heersende leefomstandigheden, terwijl andere juist zwakker zullen zijn. Deze variaties zijn dus blind, omdat zowel betere als slechtere eigenschappen ontstaan. De selectie gebeurt echter niet volledig toevallig : het is immers het milieu of de omgeving die de best passende individuen de maximale kans op voortbestaan geeft. Dit proces heeft zijn extreme vorm in wat Darwin de "artificiële selectie" noemde (de veredeling van planten en dieren). In dit proces laat de mens de natuurlijke variaties hun gang gaan, doch neemt de mens de selectie zelf over. Na elke generatie van bijvoorbeeld koeien, selecteert de mens die koeien die bijvoorbeeld het meeste melk leveren. Door deze artificiële selectie (of veredeling) onstaan mettertijd (doch veel sneller dan door omgevingsselectie zou kunnen gebeuren) koeien die alsmaar meer melk leveren. Door deze versnelling van de natuurlijke selectie is de mens erin geslaagd heel wat planten en dieren aan te passen aan de menselijke omgeving (denk maar aan alle groenten, fruit, landbouwgewassen en huisdieren).
Wat Darwin en zijn tijdgenoten nog niet wisten, omdat de wetenschap dit slechts de laatste decenia begint te ontdekken, is dat vele variaties in de natuur niet louter toevallig zijn. Bepaalde variaties blijken bevoordeeld te zijn ten opzichte van andere, door een proces van "zelf-organisatie". Bepaalde variaties krijgen daardoor een hogere waarschijnlijkheid om voor te komen, waardoor de kans op selectie ook hoger is. Deze variaties zijn als het ware geïnformeerd, terwijl de selectie in principe blind kan verlopen.
Aangezien de meeste ecosystemen zeer complex zijn, en natuurlijke selectie en zelforganisatie elkaar continu kunnen beïnvloeden en opvolgen, is het in de praktijk zelden mogelijk de effecten van beide duidelijk te onderscheiden. Vandaar dat het gezamelijke proces "zelforganisatie en natuurlijke selectie" wordt genoemd. Door de niet-lineaire en zelfversnellende processen eigen aan de zelforganisatie, leidt dit in sommige gevallen tot een "survival of the first", terwijl de natuurlijke selectie veelal gekenmerkt wordt door een "survival of the fittest". Het gezamelijke proces ligt aan de basis van de complexe fauna en flora zoals we die op aarde aantreffen, inclusief het ontstaan van de mens.
De mens is door zijn bewuste manier van denken in staat om geïnformeerde variaties zeer doeltreffend te combineren met geïnformeerde selecties, in proces dat ik verder de "artificiële constructie" zal noemen. De mens creëert artificiële variaties door bijvoorbeeld het uitvoeren van experimenten : in deze experimenten worden bepaalde factoren wel, en andere niet gevarieerd. De variaties zijn dus duidelijk geïnformeerd. Bovendien selecteert de mens het gewenste proces of product, op basis van selectiecriteria die veelal voor het uitvoering van het experiment zijn vastgelegd. Ook de selectie is dus geïnformeerd. Dit proces van artificiële constructie heeft geleid tot het onnoembare aantal "artefacten" die door de mens worden gebruikt, gaande van speren en bogen bij de primitievere mensen, tot de raketten om de ruimte te veroveren.Omdat beide stappen, zowel de variaties als de selecties doelbewust gemanipuleerd en geïnformeerd zijn, is de mens in staat om zeer complexe en effectieve constructies te bouwen.
Merk echter op de verbazingwekkende doeltreffendheid en creativiteit van bepaalde systemen in de natuur onrechtstreeks ook het gevolg is van het geïnformeerd zijn van de 2 evolutiestappen. Door de zelforganisatie is immers de variatiestap min of meer geïnformeerd, terwijl de selectiestap door een proces van natuurlijke selectie min of meer geïnformeerd verloopt. Een verschil met de artificiële constructie is wel dat de informatie in de 2 stappen niet echt is aangepast aan een lange-termijn doelstelling, zoals de mens dit wel kan doen.Het is intussen duidelijk dat het begrip "informatie" in deze evolutiecontext een ruimere betekenis krijgt dan deze van de gebruikelijke omgangstaal.
Terug naar de index van dit hoofdstuk
2.4. Wat is informatie binnen de evolutiecontext ?
Op deze vraag wat informatie is in de evolutiecontext, kunnen we ons antwoord starten met een korte definitie :
Informatie is datgene wat betekenis heeft, binnen een bepaalde context.
De betekenis van informatie voor de evolutie is dat het de selectie-waarschijnlijkheid van bepaalde evolutiewegen doet toenemen en daardoor de verspreiding beperkt.
Dit is een heel korte, doch ook heel krachtige definitie van informatie. Ze gaat immers een stuk verder dan het begrip informatie zoals we dit in onze dagelijkse omgangstaal gebruiken. In onze dagelijkse omgangstaal doet het begrip informatie ons denken aan bijvoorbeeld :
- het journaal op de televisie : hier worden nieuwe feiten aangebracht, met beelden, taal, enz.
- een mondeling gesprek : persoon x vertelt iets nieuws aan persoon y.
Kenmerkend aan beide van bovenstaande voorbeelden is dat ze uitgaan van een sterk "antroposofische visie" op informatie (een visie waarbij de mens centraal staat). Om het begrip informatie te kunnen gebruiken in onze evolutietheorie, moeten we het ontdoen van zijn menselijk karakter. Net zoals Copernicus - om een juister beeld te krijgen op het universum - de stelling moest ontkrachten dat de aarde (en dus de mens) in het centrum ligt van het universum, moeten we hier de stelling ontkrachten dat de mens centraal staat in het begrip informatie.
Informatie is datgene wat betekenis heeft, niet enkel in aanwezigheid van mensen, doch in elke context waar de betekenis de waarschijnlijkheid van een evolutierichting verandert. We kunnen de informatie dus herkennen aan het feit dat de onzekerheid in evoluties wijzigt, en de waarschijnlijkheid van evolutiewegen beinvloedt.
In zijn meest elementaire vorm is de betekenis van informatie "Kom hier" of "Blijf hier weg" (denk maar aan de rode vlag bij de stier). Informatie trekt aan of stoot af; het is een "links" of een "rechts", een "ja" of een "nee", een "nul" of een "één". Vandaar ook dat de éénheid van informatie een "bit" wordt genoemd. Dit is de afkorting van "binary digit", of "tweetallig teken", met andere woorden een bit is een "nul" of een "één". Het aantal bits of de informatie-inhoud in het voorbeeld met de stier is 3 bit. De stier moet immers 3-maal beslissen over "links" of "rechts" ; er zijn 3 opeenvolgende tweesprongen.
Het tekort aan informatie om de evolutie volledig vast te leggen is dan ook 3 bit. Het gebruik van een grote vlag vermindert echter het informatietekort tot nul : immers, bij elke tweesprong zeggen we de stier welke richting hij moet nemen. Als we enkel bij de eerste tweesprong een vlag zouden plaatsen, zou het informatietekort twee bit zijn. Met andere woorden, naarmate het informatiekort toeneemt, neemt ook de verspreiding toe en ook de onzekerheid over de uitkomst van de evolutie.
Vandaar het statement dat de betekenis van informatie voor de evolutie is dat het de selectie-waarschijnlijkheid van bepaalde evolutiewegen doet toenemen en daardoor de verspreiding beperkt.
Dit geldt zowel voor de mens, als voor een stier, als voor een inktdruppel in een glas water, als voor een electron in een computerchip, als voor elk elementair deeltje in de evolutie. In essentie is er niks menselijk aan het begrip informatie.
Terug naar de index van dit hoofdstuk
Ga naar het vervolg op de volgende pagina