Gamma attractoren in de (zelf)replicerende stroom
In wat voorafging hebben we aangetoond dat de aanwezigheid van informatie in Alfa-en Beta-attractoren de energie- en materiestroom sterk kan beinvloeden. Er ontstaan duidelijk zichtbare structuren in zowel de tijd als in de ruimte, met min of meer complexe symmetrieën.
In wat volgt zullen we dieper ingaan op de meest complexe systemen die we rondom ons kunnen waarnemen, namelijk de levende systemen. Er bestaan vele theorieën over de essentiële kenmerken van levende systemen. In de achtiende eeuw parafraseerde Immanuel Kant op ondubbelzinnige wijze : "levende organismen zijn zelforganiserende systemen bij dewelke de onderdelen als middel en als doel bestaan. Organismen zijn de mechanismen die het geheel laten bestaan, en het geheel is zodanig geordend, dat de onderdelen kunnen blijven voortbestaan."
We zullen in dit hoofdstuk levende systemen beschrijven op een wijze die ruimer is dan deze gangbaar in de klassieke biologie, omdat de beschrijvingswijze niet enkel moet geldig zijn voor de evolutie van levende systemen zoals we die nu kennen op aarde, doch voor elk mogelijk pad in de gemiddelde evolutie. Aan de basis van deze zeer complexe systemen ligt zeer complexe informatie. Opnieuw is informatie de sleutel tot de verregaande complexiteit, en tot het behoud van de "ver-van-evenwichtssituatie" die kenmerkend is voor de Beta-systemen en Gamma-systemen.
Het grote onderscheid tussen Beta-attractoren en Gamma-attractoren is de complexiteit van informatie. Waar bij de Beta-systemen, de informatie gevormd wordt door kenmerken van het proces zelf, is bij de Gamma-attractoren de informatie gekristalliseerd in complexe (meestal symbolische) gecodeerde vorm. Bij de ontwikkeling van de het levend systeem, moet de gecodeerde informatie vertaald worden in de vorming van een nieuw levend systeem. Dit betekent dat de Gamma-attractor ook de "infrastructuur" in zich houdt om zowel de vertaling van de complexe informatie te kunnen doen, als om de opbouw van het levend systeem te verwezenlijken. Kenmerkend hierbij is dat zowel de hardware (M/E) als de software (I) van de attractor perfect op elkaar zijn afgestemd : ze zijn complementair. Daardoor kan het systeem zichzelf uiteindelijk repliceren op basis van de ontwikkelings-instructies. Dit is het meest kenmerkende aspect van het leven.
Levende systemen zijn te vergelijken met machines die instructies bevatten om copies van zichzelf te produceren, en die deze instructies ook in realiteit kunnen omzetten. Deze vergelijking met machines of menselijke artefacten laat duidelijk zien dat de meeste - door de mens gebouwde machines of artefacten - hier ver van afstaan. De meeste machines verschillen zeer sterk van de producten die ze kunnen produceren ; dit geldt zowel voor "hardware"-producerende als voor "software"-producerende artefacten.
In dit hoofstuk zullen we eerst aantonen waarin het voornaamste onderscheid bestaat tussen de Gamma-attractoren en de andere attractoren die we reeds besproken hebben.
Daarna zullen we dieper ingaan op het ontstaan van "Gamma-attractoren" en "Gamma-replicatoren" op onze aarde.
Uiteindelijk zullen we een model beschrijven van wat een "universele Gamma-attractor" zou kunnen genoemd worden.Terug naar de index van dit hoofdstuk
1. Gamma-attractoren en Gamma-replicatoren
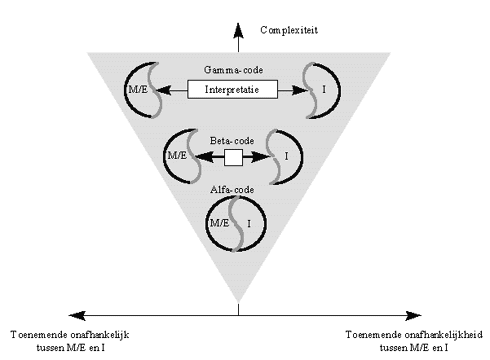
Het onderscheid tussen Alfa-, Beta- en Gamma-informatie ligt essentieel in de mate waarin er een directe relatie is tussen de drager van de informatie, en de betekenis ervan.
Figuur H7F2 : relatie tussen de informatie en de drager ervan bij Alfa-, Beta- en Gamma-informatie
Bij de Alfa-attractoren is de relatie tussen de de materiële drager, en de betekenis van de informatie zeer direct. De Alfa-informatie is daarom in veel gevallen niet of nauwelijks voor interpretatie vatbaar. De herhaalbaarheid, en dus ook de voorspelbaarheid van Alfa-interacties is daarom ook hoog, met als gevolg dat ze veelal in mathematische wetmatigheden te beschrijven zijn.
Bij Beta-informatie is dit minder het geval, omdat de betekenis het gevolg is van een dynamisch proces, dat zelf onderhevig is aan talrijke variabelen, die de stabiliteit van de interpretatie beïnvloeden.Bij Gamma-informatie echter is de codering symbolisch, en heeft de code op zich geen enkele fysische betekenis meer. De betekenis is daarom volledig afhankelijk van de interpratie die de interactor er binnen de gegeven context aan geeft ; met andere woorden, de betekenis is volledig conventioneel.
Het was de Amerikaanse wetenschapper Charles Sanders Peirce (1838-1914) die "het proces van betekenisgeven" of het "semiotisch proces" voor het eerst als een drieluik of "triade" voorstelde (zie onderstaande figuur links). Hij zette zich hiermee af tegen de school van Saussure, die informatie voorstelde als een tweeluik, met aan de ene kant de drager, en aan de andere kant betekenis ervan.
Uit het bovenstaande blijkt dat in het geval van Alfa-informatie, de ruimte voor interpretatie zo nauw is, dat de "diadische" voorstelling van Saussure toereikend is : bij Alfa-informatie is immers nauwelijks ruimte voor interpretatie. Dit geldt echter niet voor de Gamma-informatie, terwijl de Beta-informatie ergens tussenin zweeft, doch eerder aan de diadische kant.
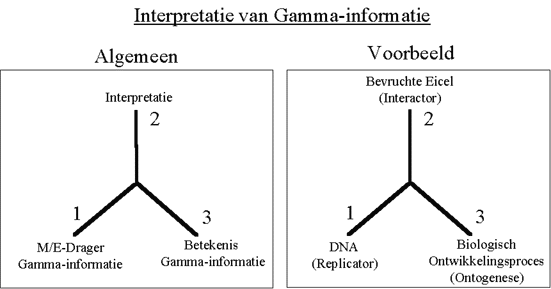
Figuur H7F3 : Het informatieverwerkingsproces volgens Peirce, toegepast op Gamma-informatie van een biologisch organisme
Nemen we als voorbeeld van Gamma-informatie het DNA van een biologisch organisme. Dit DNA bevat een beschrijving van het organisme, en laat het organisme toe zichzelf te repliceren. We zullen het daarom verder de replicator noemen.
Uit het DNA op zich, zal echter nooit een levend organisme ontstaan. De genetische code moet immers eerst geïnterpreteerd worden door de bevruchte eicel. Pas als gevolg van deze interpretatie start het biologisch ontwikkelingsproces (de "ontogenese").
De bevruchte eicel is essentieel in dit proces, want enkel deze eicel kan de code lezen, en omzetten in een organisme. De genetische code is datgene wat van de ene generatie op de andere wordt overgedragen en bevat enkel een beeld of replica van het levend systeem.
Omwille van het belang van de interpretatie van de Gamma-informatie door de betrokken interactor, zullen we aannemen dat de Gamma-attractor zowel de ontwikkelingsinformatie uit het DNA bevat, als de informatie voor de interpretatie ervan. Zonder het ontwikkelingsmechanisme, krijgt de Gamma-informatie immers nooit zijn volle betekenis.
Men kan zich hier de vraag stellen waarom zich in de natuur, en nog meer de gemiddelde evolutie, dergelijke complexe informatieprocessen zouden ontwikkelen.
Het antwoord hierop is dat de symbolische informatie over een competitief voordeel beschikt, dat niet te evenaren is door de Alfa- en de Beta-informatie, namelijk de "dubbele articulatie".
De menselijke taal is ook een symbolische taal, en dus gekenmerkt door dubbele articulatie : we gebruiken woorden (het eerste niveau van articulatie), en elk woord heeft op zich een betekenis. We combineren deze woorden echter tot zinnen (het tweede niveau van articulatie), waarmee we om het even welk object of abstracte gebeurtenis tot in het kleinste detail kunnen beschrijven. Dubbele articulatie biedt dus een enorme hefboom voor het toenemen van de complexiteit in de evolutie. Bovendien laat het toe om niet enkel andere objecten -buiten het systeem- te beschrijven, doch ook het systeem zelf. Dit is wat het DNA doet in biologische organismen, met de overleving tot gevolg over de verschillende generaties. Hetzelfde principe laat de mens ook toe over zichzelf na te denken en zichzelf te beschrijven, met andere woorden het is dus ook de basis van het menselijke bewustzijn en de culturele overlevering.Sommige soorten ééncellige organismen planten zich voort door kloneren of a-sexuele voortplanting ; dit is de eerste vorm van voortplanting die op onze aarde is ontstaan bij levende organismen.
Elk individu (interactor) van de populatie van deze bacterie is identiek aan alle andere klonen (de fouten door het maken van copies worden hier voorlopig verwaarloosd).
In dit geval is de Gamma-attractor voor de soort, identiek aan de Gamma-attractor voor het organisme.
Later in de evolutie is echter sexuele voortplanting ontstaan, waarbij bij elke paring een recombinatie van het genetisch materiaal gebeurt. Hierdoor is elk individu verschillend van het andere. Toch zijn de verschillende individuen van de soort zeer gelijkend en hebben de individuen een scala aan gemeenschappelijke kenmerken. In dit geval is het niet zeer zinvol om te spreken van een Gamma-attractor op het niveau van het individu, omdat er geen twee gelijke individuen zijn. De Gamma-attractor situeert zich in dit geval op het niveau van de soort van levende systemen.
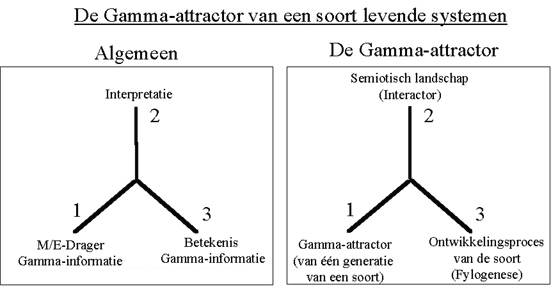
Figuur H7F4 : Het informatieverwerkingsproces volgens Peirce, toegepast op Gamma-informatie van een soort levende systemen
De populatie van individuen in de soort, vertoont hierbij een collectief gedrag, dat aangepast is aan de (semiotische) omgeving. Dit "gemiddeld collectief gedrag" van de populatie wordt hierbij bepaald door de Gamma-attractor van de populatie.
In dit geval is het de omgeving die door het uitwisselen van signalen, de kenmerken van de Gamma-attractor bijstuurt, en zo de ontwikkeling van de soort bepaalt (de fylogenese). De interpretatie is hierbij een interactief proces tussen de populatie en zijn omgeving (geen éénwegsproces).
Het proces van betekenis-vorming zoals door Peirce beschreven impliceert een zekere volgorde in de evolutie. Bestaande informatie wordt geïnterpreteerd, en hierbij omgezet in nieuwe fysische structuren. De omgeving voert door deze interpretatie een zekere natuurlijke selectie uit.Deze volgorde sluit ook de theorie van Lamarck uit.
Het Lamarckisme was een evolutietheorie die populair was voor het Darwinisme. Ze ging er precies van uit dat wijzigingen in de M/E structuur van het levend systeem, automatisch werden overgedragen naar de volgende generatie. De lange nek van een giraf is volgens deze theorie ontstaan omdat de giraffen steeds hoger moesten grijpen naar de bladeren in de bomen, waardoor hun nek werd uitgerokken. Volgens het Lamarckisme werden zo de nekken van opeenvolgende generaties steeds langer. Het Lamarckisme komt overeen met een proces dat we kunnen kenmerken met de boutade "van atoom tot bit".De werkelijke evolutie is net omgekeerd. Door toevallige mutaties in de genetische informatie van de giraffen, moeten giraffen zijn ontstaan met een nek die iets langer was dan die van de concurrentie. Daardoor konden zij bij voedselschaarste langer overleven, en de overhand krijgen in de populatie : de Daardoor konden "de lange nekken" ook meer paren, en kregen ze meer nakomelingen : de "lange nekken" werden dus geselecteerd uit een bestaande informatiepool.
In elke nieuwe generatie zaten wel enkele giraffen met langere nekken, waardoor na vele generaties, onder de selectiedruk van voedselschaarste, een populatie onstond van giraffen met lange nekken zoals we die nu kennen.Hetzelfde evolutiemechanisme geldt voor alle levende systemen gebaseerd op Gamma-attractoren. De omgeving (het semiotische landschap) selecteert ad random mutaties in de Gamma-informatie.
Dit proces hebben we eerder gekarakteriseerd als "’it and fit from bit" (vanwaar ook de titel "van bit tot atoom en ecosysteem").We zullen in een volgend hoofdstuk zien dat dit semiotisch landschap het ecosysteem en zijn populaties doet evolueren in de richting van fluxmaximalisatie, naarmate het meer matuur wordt.
In dit hoofdstuk zullen we echter eerst het ontstaan van de eerste replicatoren op aarde nader bekijken, om daarna op zoek te gaan naar de kenmerken van een "universele Gamma-attractor".
Terug naar de index van dit hoofdstuk
2. Het ontstaan van de eerste Gamma-replicatoren
Wanneer we het onstaan van de eerste Gamma-replicatoren op aarde willen nagaan, moeten we het ontstaan van het eerste leven op aarde van dichterbij gaan bekijken.
Zoals reeds beschreven in het eerste hoofdstuk, is het leven op aarde vrij snel ontstaan. Onze aarde is ongeveer 4.6 miljard jaar oud, en ongeveer één miljard jaar later, toen uit de oceanen het eerste sedimentair gesteente was gevormd, ontstond wellicht ook het eerste leven op aarde. Fossiele overblijfselen van organisch materiaal afkomstig van levende cellen zijn ontdekt bij electronenmicroscopische studie van rotsgesteenten uit de precambrische periode.
Eén van de meest verspreide theorieën over het leven op aarde is deze van de Russische chemicus Aleksandr Oparin, die in 1924 suggereerde dat - als gevolg van een reeks niet-biologische reactiestappen - er zich organische molecules van steeds grotere complexiteit hebben gevormd. Hieruit ontstonden uiteindelijk molecules met een zelf-replicerend vermogen en uiteindelijk ook levende systemen. Naarmate deze zich meer verspreiden, wijzigde de aardatmosfeer zich in een atmosfeer die steeds gunstiger werd voor de biologische evolutie.
De atmosfeer van de aarde was het eerste miljard jaar te vergelijken met de huidige atmosfeer van Jupiter, Saturnus, Uranus en Neptunus. Rond deze planeten hingen -voor de mens - giftige wolken van methaan, ammoniak, waterdamp en waterstof. De atmosfeer bevatte weinig zuurstof. In die tijd echter was de atmosfeer meer doorlatend voor UV-stralen en kosmische stralen uit de ruimte. Door de opbouw van statische electriciteit, zorgden bliksemschichten voor energetische schokgolven die complexe chemische reacties konden versnellen. Daardoor werd de aarde steeds rijker aan koolstofmonoxide en koolstofdioxide (nodig voor de opbouw van levende cellen).
Deze periode was gunstig voor de "chemische evolutie" op aarde. In deze periode werden de bouwstenen gevormd die nodig zijn voor het ontstaan van leven zoals we dit op aarde kennen.
Geleidelijk aan koelde de aarde af, en werd de electrische activiteit geringer. Na het ontstaan van het eerste leven, ontstonden ook cellen die door photosynthese de aardatmosfeer hebben aangereikt met het zuurstof. Zo werd de aarde uiteindelijk gunstig voor het onstaan van de biodiversiteit die we nu kunnen waarnemen.
We zullen nu de ontwikkeling van levende wezens eens meer in detail bekijken.
Terug naar de index van dit hoofdstuk
2.1. De vorming van organische bestanddelen
Voor de eerste stap van de vorming van levende systemen op aarde was er meer nodig dan de anorganische molecules die bij de condensatie van onze planeet zijn ontstaan. De basismolecules van de levende cellen werden gevormd door organische verbindingen (verbindingen met koolstof, in combinaties met voornamelijk waterstof, zuurstof en stikstof).
Omdat deze elementen in de primitieve atmosfeer overvloedig aanwezig waren, tesamen met grote hoeveelheden energie, waren de omstandigheden van de primitieve atmosfeer gunstig voor organische syntheses, die in onze huidige atmosfeer nog nauwelijks buiten levende cellen voorkomen.
In de toendertijd vooruitstrevende experimenten van Stanley Miller in 1953, werden de atmosferische omstandigheden van de primitieve atmosfeer in labo-omstandigheden gereproduceerd. Hij ontdekte dat electrische ontladingen in een mengsel van methaan, ammoniak (een bron van stikstof), water en waterstof, resulteerden in de vorming van aminozuren. Met zijn experiment had hij een essentiële stap in de evolutie van het leven, namelijk de vorming van organische uit anorgische bestanddelen, experimenteel bewezen.
Naast de spontane vorming op aarde, bereiken organische molecules de aarde ook vanuit de ruimte. Sporen van organische bestanddelen zijn immers aangetroffen in meteoren en kometen. Dit wijst erop dat de vorming van voor het leven essentiële bestanddelen ook buiten de aarde niet uitzonderlijk is.
Vanwaar de eerste organische molecules ook afkomstig waren, ze vormden in hun waterig milieu de pre-biotische oersoep die nodig was voor verdere ontwikkeling van het leven.
Terug naar de index van dit hoofdstuk
2.2. De vorming van zelf-replicerende polymeren
Een belangrijk kenmerk van levende systemen is dat ze de gecodeerde instructies (de informatie I) bevatten om hun eigen structuren (M/E) op te bouwen en te repliceren.
Sinds jaren is een geaccepteerde theorie voor het onstaan van het leven gebaseerd op het ontstaan van zelfreplicerende molecules (RNA, DNA), uit de prebiotische oersoep.
We kunnen ons eerst algemeen de vraag stellen welke kenmerken een molecule moet hebben, opdat ze in staat zou zijn haar eigen synthese te bevorderen.
Deze vraag is de hamvraag voor academici die in laboratoria kunstmatige zelfreplicerende mocules trachten te bouwen.
Julius Rebek is één van de academici (verbonden aan het MIT als hoogleraar chemie) die een antwoord tracht te formuleren op deze vraag.
"Molecules, natuurlijk of synthetisch, zijn in staat tot zelfreplicatie als hun vorm en chemie een eigenschap hebben die complementariteit wordt genoemd". Door de specifieke vorm kan de molecule atomen of groepen van atomen aantrekken en dicht bij elkaar brengen. De kans op een nieuwe chemische binding hangt niet enkel af van een passende ruimtelijke structuur, doch ook van de aangepaste chemische bindingen die de atomen of molecules samenhouden in groepen of complexen. Deze groepen of complexen bestaan dikwijls slechts gedurende heel korte tijd (microseconden of nanoseconden), doch voldoende lang om een echte chemische binding te veroorzaken.
Van zodra deze complexen gevormd zijn, beschermen de moleculaire oppervlakken elkaar, waardoor het moeilijker wordt voor destructieve solventen (zuren, basen, oxidantia) om de partners van elkaar te verwijderen. Op deze manier is er voldoende tijd om de sterkere (covalente) bindingen tot stand te brengen tussen de complementaire molecules.
Dergelijke manier van koppeling voor replicatie wordt gebruikt door het DNA.
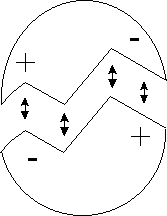
Een éénvoudig voorbeeld van complementariteit is de aanwezigheid van concave en convexe oppervlakten (zie onderstaande figuur). Een concaaf moleculair oppervlak kan zijn convexe tegenpool herkennen en omgeven. Bovendien kan het als matrijs fungeren voor assemblage van de convexe molecule vanuit zijn samenstellende componenenten. Dit is dus het "sleutel op het slot"-principe dat kenmerkend is voor de Alfa-informatie. Alfa-informatie speelt dus een belangrijke rol in de vorming van Gamma-informatie die nodig is voor zelfreplicatie.
Figuur H7F5 : Twee complementaire molecules herkennen elkaar omdat hun ruimtelijke en chemische structuur aan elkaar is aangepast. De plus- en mintekens wijzen op electrostatische krachten.
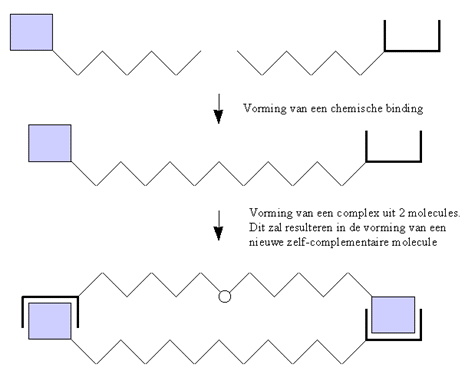
Een ander mogelijk scenario voor zelfreplicatie is voorgesteld in onderstaande grafiek. Twee complementaire molecules kunnen met elkaar binden op een plaats die verschillend is van de herkenningsplaats (in het vorige scenario viel de binding op de herkenningsplaats). Ze vormen één enkele molecule , waarvan het éne eind complementair is aan het andere eind, en de hele molecule complementair is met zichzelf.
De molecule draagt dus zowel de informatie als het mechanisme in zich om zichzelf te repliceren.
Figuur H7F6 : zelfreplicatie ontstaat wanneer twee complementaire molecules samenkomen en een zelfcomplementaire molecule vormen. Deze molecule verzamelt dan copies van de originele componenten en brengt ze samen, zodat ze kunnen reageren. Het resultaat is een copie van zichzelf.
Terug naar de index van dit hoofdstuk
2.3. Hypercycli en de vorming van zelfreplicerende polymeren
Bovenstaand algemeen mechanisme toont aan hoe complementariteit (als bijzondere vorm van zelfreferentie) een basisprincipe vormt bij het ontstaan van zelfreplicerende molecules. Om de vorming van lange ketens van meerdere molecules op een foutloze en reproduceerbare wijze te laten verlopen, is complementariteit op zichzelf niet toereikend.
Nobelprijswinnaar Manfred Eigen heeft een zelforganizerend mechanisme voorgesteld, dat de ontwikkeling van complementaire I- en M/E- structuren met toenemende complexiteit kan verklaren. Het principe stelt dat een "zelf-reproducerende hypercyclus" wordt gevormd. Deze evolutie kan doorgaan in een niet-evenwichtstoestand, waarbij het open systeem continu wordt gevoed door infllux van energie, ter compensatie van de entropieproductie.
Het model is in onderstaande figuur samengevat.
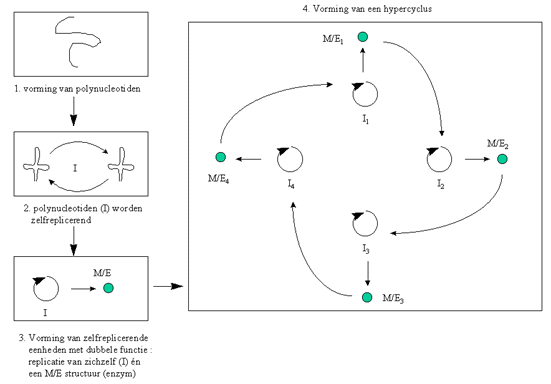
Figuur H7F7 : een zelfproducerende hypercyclus (naar M. Eigen & P. Schuster, 1979)
Stap 1 : vorming van polynucleotiden
In een eerste fase van de evolutie worden ketens gevormd van nucleotiden, met name polynucleotiden.
De vorming van polynucleotiden is nog te verklaren door een mechanisme zoals dit door Stanley Miller is ontwikkeld. Voor de verdere stappen in de ontwikkeling zullen we echter beroep moeten doen op andere ontwikkelingsmechanismen. De logica achter deze mechanismen moet echter rekening houden met de beperking dat slechts 2 factoren invloed kunnen hebben :
- de natuurlijke overvloed aan componenten
- de sterkte van interacties is volledig bepaald door de aard en vorm van de molecules (enkel Alfa-informatie aanwezig)
Wat de overvloed aan componenten betreft, werd experimenteel aangetoond dat nucleïnezuurachtige bestanddelen (de enige bekende natuurlijke molecules die aanleiding geven tot zelfreplicatie) inderdaad in voldoende mate gevormd kunnen worden. Het gaat hier meer specifiek over de nucleïnezuren Adenine, Uracil, Citosine en Guanine. De vorming van de meer energierijke nucleotiden, en in het bijzonder hun energierijke fosfaatvormen, is minder vanzelfsprekend via de pre-biotische mechanismen, doch is niet onmogelijk. Dit is niet onbelangrijk, omdat het voornamelijk deze energierijke nucleotiden zijn die in aanmerking komen voor polymerisatie.
De mate waarin polymeren gevormd worden, is afhankelijk van de concentratie waarin de monomeren (enkelvoudige molecules) voorkomen.
De polynucleotiden hebben de eigenschap zich "te kronkelen" en een driedimensionele structuur te vormen, op zodanige wijze dat de potentiële energie binnenin de molecules minimaal wordt. Deze "secundaire structuur" wordt enkel en alleen bepaald door de volgorde van de nucleotiden, en de cruciale eigenschap van de nucleïnezuren om zich paarsgewijs te koppelen in DNA :
- adinine koppelt steeds met thymine (of uracil in RNA) : hierbij wordt het "basepaar" A-T (of A-U) gevormd
- guanine koppelt steeds met cytosine : hierbij wordt het "basepaar" G-C gevormd
Deze spontane koppeling is de basis voor de vorming van zelfreplicerende complexen, zoals dit ook in de voorbeelden in voorgaande paragraaf is aangetoond.
Stap 2 : vorming van zelfreplicerende polynucleotiden
Tussen de grote hoeveelheden polynucleotiden, ontstaan er door toeval enkele "witte raven", die zelfreproducerende segmenten bevatten. In deze segmenten is de ruimtelijke structuur zodanig, dat de koppeling van baseparen optimaal verloopt. Dit betekent dat een dergelijk segment (bijvoorbeeld met drie nucleotiden A-C-G), zodanig gevormd is, dat zich bij de A een U (of T) koppelt, bij de C een G. Bovendien bevordert de structuur een binding tussen de 2 bijeenliggende molecules. Hetzelfde gebeurt met de derde molecule.
Dit nieuwe nucleotide met 3 nucleïnezuren is nu een spiegelbeeld (complementaire structuur) van het origineel. Als dit spiegelbeeld nu ook zelfreplicerend is, krijgen we een copie van het originele minipolymeer.
Uiteindelijk ontstaan uit de polynucleotiden met zelfreplicerende segmenten, polynucleotiden die volledig zelfreplicerend zijn. De kans dat dit gebeurt is zeer klein maar bestaat. Van zodra dit echter gebeurt, heeft dit polynucleotide een "competitief voordeel" ten overstaan van de niet zelfreplicerende polynucleotides. Daardoor zal het niet enkel toenemen in aantal, het zelfreplicerende karakter zal ook steeds beter worden. Deze optimalisaties gebeuren op een wijze die te vergelijken is met de Darwinistische natuurlijke selectie. Door toevallige fouten bij de zelfreplicaties zullen zelfreplicerende molecules ontstaan die iets beter dan de andere de nucleïnezuren binden. Hierdoor zullen ze zich sneller repliceren dan andere, waardoor ze uiteindelijk de overhand zullen uitmaken in de populatie van polynucleotiden. Deze natuurlijke selectie gaat door, tot een "optimale vorm" is bekomen.
Stap 3 : vorming van zelfreplicerende molecules met dubbele functie.
Onder deze "geoptimaliseerde" zelfreplicerende molecules, zullen opnieuw enkele "witte raven" voorkomen. Er zullen "altruïstische" molecules ontstaan, die niet enkel replicatie van zichzelf mogelijk maken, doch die ook de vorming van een andere molecule versnellen. Dergelijke reactieversnellende molecules worden in de biochemie enzymen genoemd.
Door natuurlijke selectie zullen meer en meer zelfreplicerende molecules met enzymatische functie ontstaan. De ruimtelijke structuur van de molecules wordt hierbij steeds belangrijker : het zijn immers de specifieke concave en convexe ruimtes binnenin de molecules die de enzymatische functie veroorzaken.
Hierbij ontstaan bijvoorbeeld molecules gespecialiseerd in het aanbrengen van reeksen van nucleotiden. Kenmerkend voor de evolutie op aarde is dat er een type molecules is ontstaan gespecialiseerd in het aanbrengen van 3 aminozuren (deze worden de transfer-RNA’s genoemd).
De meer en meer complex wordende molecules, met niet enkel een secundaire maar zelfs tertiaire structuur, behoren tot de klasse van wat we in de biochemie de eiwitten noemen. In onze complexer wordende oersoep komen dus naast complexere polynucleotiden ook eiwitten voor.
Stap 4 : de vorming van hypercycli
Naarmate de verscheidenheid aan pre-biotische molecules toeneemt, beginnen ook de interacties toe te nemen. Hierdoor onstaan de hypercycli.
Een dergelijke hypercyclus bestaat uit een aantal zelfreplicerende molecules, die "in een kringloop " werken. De enzymatische functie van het eerste polynucleotide versnelt de vorming van het tweede zelfreplicerende molecule, dat op zijn beurt de vorming van een derde versnelt enz. Het laatste in de rij versnelt uiteindelijk de vorming van het eerste zelfreplicerende molecule.
Deze hypercyclus bevat dus twee niveau’s van terugkoppeling (nog een vorm van zelfreferentie) :
- ten eerste bevorderen de molecules hun eigen replicatie
- ten tweede bevorderen ze in kring elkaars replicatie.
Kenmerkend voor deze hypercyclus is een zelforganiserende dynamiek, gelijkaardig met de zelforganiserende processen die beschreven zijn bij de Beta-attractoren. De cyclus is op zichzelf een dissipatieve structuur, met een aantal voordelen voor het evolutieproces :
- door de terugkoppeling ontstaat een soort controlerende functie, die toelaat polynucleotiden met grotere informatie-inhoud te maken ;
- de vorming van een complex repliceerapparaat wordt mogelijk : hierbij kan de dubbele functie van de enzymes stilaan vervallen : voor elke functie kunnen gespecialiseerde enzymes ontstaan ;
- de betrouwbaarheid van de replicaties verhoogt : er worden minder fouten gemaakt.
Terug naar de index van dit hoofdstuk
2.4. De oorsprong van een complex metabolisme
De biologie is in zijn geheel gekenmerkt door talrijke complexe systemen.
Voorbeelden hiervan zijn de duizenden genen die elkaars werking beïnvloeden ; het netwerk van cellen en molecules van het immuunsysteem ; de ontelbare neuronen die met hun neurale netwerken aan de basis liggen van complex gedrag en het ecosysteem als semiotisch landschap en voedselweb voor de co-evolutie van verschillende levende systemen.De theorie van Eigen en Schuster over het ontstaan van zelfreplicerende mechanisme levert een vrij goed antwoord op de vraag hoe zelfreplicatie kan ontstaan. We moeten ons echter ook afvragen hoe, samen met de zelfreplicatie, de complexiteit van het biologische metabolisme is ontstaan. Het metabolisme van een (biologisch) levend systeem is uiterst complex. Het bestaat uit een zeer uitgebreid netwerk van chemische transformaties, waarvan sommigen de orde verhogen, en anderen de orde verlagen. Elk van deze transformaties is ook te beschouwen als een stroom van materie, energie en informatie.
Recent onderzoek op basis van computermodellen laat uitwijzen dat het metabolisme een netwerk zou kunnen zijn dat als een Beta-proces door zelforganisatie is ontstaan.
Eén van de opinieleiders in dit verband is Stuart Kauffman, van het befaamde Santa Fe instituut in de verenigde staten. Kauffman heeft aangetoond dat vele complexe kenmerken van levende systemen zijn ontstaan door een proces van spontane zelforganisatie, dat hand in hand gaat met het proces van natuurlijke selectie zoals dit door Darwin is beschreven. De combinatie van beide evolutie-principes heeft hij "zelf-organisatie en selectie" genoemd.
Sinds Darwin hebben vele biologen natuurlijke selectie als vrijwel de enige bron van orde in levende systemen gezien. Darwin kon echter onmogelijk de principes van de complexe Beta-processen kennen, zoals deze de laatste decennia zijn ontwikkeld. De grote orde van levende systemen is daarom wellicht in de eerste plaats een proces van zelforganisatie, waarop natuurlijke selectie zijn gang kon gaan. De studies die deze conclusies onderbouwen zijn nog niet afgewerkt, doch op basis van mathematische modellen van levende organismen, kunnen de kenmerken van deze organismen vrij goed voorspeld worden. Het huwelijk tussen zelforganisatie en natuurlijke selectie is dus in zijn beginfase.
Kauffman heeft kunnen aantonen dat als er een kritische hoeveelheid molecules aanwezig is, dat er een collectief gedrag ontstaat, waarbij autokatalytische eenheden van polymeren leiden tot een gezamelijk metabolisme. Voorbij een kritisch punt, gaan die verschillende sub-systemen van chemische reacties, samenwerken en een collectief metabolisme doen onstaan. Het metabolisme is als het ware de bio-chemische resulatante van het proces dat hier de fluxmaximalisatie is genoemd.
Het metabolisme is volgens Kauffman daardoor het gevolg van een spontaan collectief gedrag, dat verder verfijnd is door natuurlijke selectie.
Uit de talrijke computersimulaties die door Kauffman zijn uitgevoerd, blijkt inderdaad dat complexe netwerken de eigenschap hebben om door spontane zelforganisatie netwerken te vormen die een drastisch verhoogde coherentie vertonen. Bovendien blijkt uit de studies dat het aantal mogelijke "oplossingen" of "emergente metabolismen" binnen deze netwerken beperkt is. Ook binnen het gamma van "mogelijke" metabolismen, blijken er dus een bepaald aantal een verhoogde probabileit te vertonen : het zijn als het ware de "attractoren" van de metabolismen.
De theorie van Kauffman geeft niet enkel een verklaring voor het ontstaan van complexe metabolismen, doch ze toont ook aan dat de hypercycli die beschreven zijn door Eigen en Schuster, door eenzelfde proces van spontane emergentie, een grotere waarschijnlijkheid hebben om te ontstaan dan oorspronkelijk gedacht.
Het ontstaan van zelfreplicerende mechanismen en een metabolisch netwerk, is daarom wellicht een proces van co-evolutie als gevolg van het proces van zelforganisatie.
Terug naar de index van dit hoofdstuk
2.5. Compartimentering van zelf-replicerende eenheden
De metabolismen en replicatoren die ontstaan door zelforganisatie hebben als "attractoren" niet enkel het kenmerk van zelforganisatie (ze bevorderen hun eigen onstaan), doch ze bevorderen ook hun eigen stabiliteit (ze bevorderen hun zelfbehoud). Homeostase is immers een belangrijk kenmerk van metabolismen.
Binnen deze evolutie ontstaan ook langzaamaan eiwitten en molecules die niet zelf-reproducerend zijn, doch waarvan de reproductie ondersteund wordt door interactie met enzymes aanwezig in het milieu.
Door de hypercycli onstaat dus geleidelijk een scheiding tussen I- en M/E structuren : er ontstaan structuren gespecialiseerd in het behoud en de vertaling van de informatie, en er onstaan losstaande structuren met andere functies. De comparimentering van de functionele eenheden is wellicht ontstaan door natuurlijke selectie, om de invloed van variaties door het milieu te beperken op het steeds verfijndere netwerk van chemische reacties.
Bij virussen bijvoorbeeld is een celwand onstaan die volledig bestaat uit eiwitten. Deze eiwitten zorgen voor bescherming van de inhoud van de cel, en verzekeren dus een langere overleving.
Het ontstaan van een celwand (als begrenzing) is belangrijk voor levende wezens. Door een celwand- of celmembraan evolueert het systeem van een volledig open systeem, dat blootstaat alle alle mogelijke agressieve aspecten van het milieu, naar een "half-open systeem" dat niet enkel bescherming biedt, doch ook voor selectieve input en output van metabolische materialen. Experimenten hebben aangetoond dat membranen, gelijkaardig aan de celmembranen van primitieve ééncellige organismen, zich spontaan kunnen vormen.
Geleidelijk aan kan binnen de celwand een metabolisme ontstaan (een compleet netwerk van M/E- en I-kanalisaties), waarvan de complexiteit door natuurlijke selectie langzaam toeneemt.
- Uiteindelijk zijn op aarde zo de eerste levende wezens ontstaan.
De beschrijving hierboven is heel summier, en geeft enkel de essentie weer. Er werd een mogelijk scenario geschetst voor de evolutie van levende systemen in de biologische evolutie. Het was hierbij echter zeker niet de bedoeling een volledige historische reconstructie te maken. Conclusie uit het bovenstaande is dat het spontaan onstaan van Gamma-replicatoren, en dus van levende systemen, een realistisch scenario is. Het onstaan van zelfreplicerende eenheden is weliswaar een zeer uitzonderlijke gebeurtenis, doch het is een gebeurtenis die zich zal voordoen van zodra de omstandigheden er rijp voor zijn. Levende systemen zijn dus wellicht inherent aan de gemiddelde evolutie.
In werkelijkheid heeft dit evolutieproces tientallen tot honderden miljoenen jaren geduurd. Wetende dat de homo sapiens slechts ongeveer 300000 jaar geleden is ontstaan, is dit toch wel een relatief lange periode.Terug naar de index van dit hoofdstuk
3. Levende systemen en Gamma-attractoren
3.1. Een universele Gamma-attractor voor levende systemen ?
In de voorgaande paragrafen is aangetoond dat het onstaan van levende systemen in de evolutie een spontaan proces is, dat zich kan voordoen van zodra de omstandigheden ervoor geschikt zijn. Omdat levende systemen een wezenlijk element uitmaken van de gemiddelde evolutie, moeten we ons de vraag stellen hoe die levende systemen er dan wel "gemiddeld" zullen uitzien. Wat zijn de minimale kenmerken die eigen zijn aan een levend systeem ?
Aan het beantwoorden van deze vraag hebben al vele academici hun wetenschappelijke carrière gewijd.
In de context van dit boek kan deze vraag worden vertaald als "Wat zijn de kenmerken van een universele Gamma-attractor ?". Aangezien elk levend systeem gekenmkerkt is door heel wat contextuele (en dus toevallige) elementen, zullen we ons moeten beperken tot zeer algemene kenmerken.
Linneaus bijvoorbeeld beschreef de biologische organismen voornamelijk door de verschillen tussen de soorten op te sommen, en de soorten in klassen in te delen. Darwin ging een stap verder, en onderzocht hun evolutie. Hij kwam hierbij tot de vaststelling dat alle levende wezens op aarde moeten ontstaan zijn uit éénzelfde oersoort. Het is door de evolutie en differentiatie vanuit éénzelfde stam, dat de gelijkenissen tussen de soorten te verklaren zijn.
Een volledig ander standpunt op levende wezens krijgen we wanneer we ze benaderen vanuit de systeemtheorie en cybernetica. Deze analyse van "wat is een levend systeem" focuseert eerder op de gelijkenissen tussen de levende systemen, dan tussen de verschillen, door eerder te zoeken naar een synthese dan te streven naar een gedetailleerde analyse. Deze aanpak is dan ook beter geschikt voor het zoeken naar de "universele Gamma-attractor".Een belangrijke aanzet in dit systeemdenken over levende wezens is gegeven door Alfred North Whitehead (1861-1949), in zijn "filosofie van het organisme". Voor Whitehead was een levend wezen opgebouwd uit systemen, die op hun beurt ook uit systemen bestaan. Vanuit de vaststelling dat er binnen de fysica een hiërarchie van systemen bestaat, vertaalde hij dit naar de biologische en sociale fenomenen.
Whitehead legde hiermee de basis voor latere systeemdenkers. Zo publiceerde Von Bertalanffy in de jaren 1950 zijn "General systems theory", die een belangrijke basis vormde voor het algemene systeemdenken.
Op basis van deze ideeën, bouwde James Grier Miller - een leerling en bewonderaar van Whitehead - met een multidisciplinaire onderzoeksgroep zijn "Theorie van de levende systemen" uit. Levende systemen zijn volgens Miller systemen van materie en energie, die georganiseerd zijn door informatie. Deze levende systemen zijn open systemen, die bestaan uit verschillende hiërarchische niveau’s. Op elk niveau zijn er kenmerkende inputs, throughputs en outputs van materie, energie en informatie. Deze verwerkende processen worden uitgevoerd door subsystemen, waarvan er een aantal heel kenmerkend en essentieel zijn voor levende systemen.
Hieronder wordt een model voorgesteld dat is uitgewerkt op basis van de ideeën van Miller.
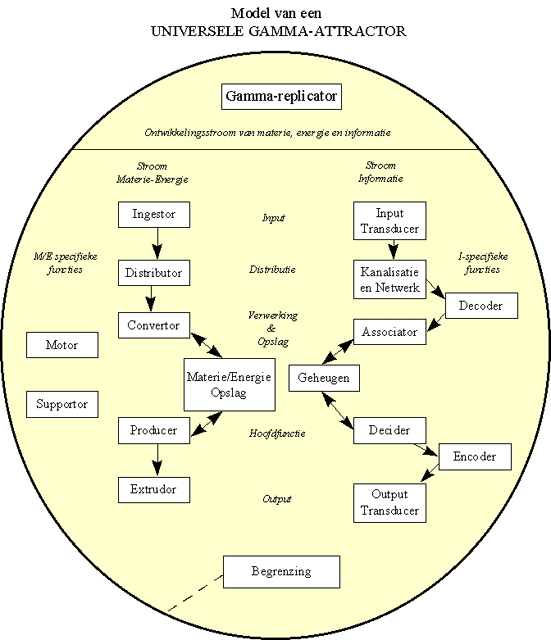
Figuur H7F8 : een model van een universele Gamma-attractor (naar een algemeen model van levende systemen volgens Miller, 1978)
Deze beschrijving van de de M/E en I-stroom doorheen een levend systeem zal binnen dit werk gesitueerd worden als de "Universele Gamma-attractor van de levende systemen" in de gemiddelde evolutie. Dit wil zeggen dat, als er zich binnen de gemiddelde evolutie levende systemen zullen ontwikkelen, dat deze zullen voldoen aan de kenmerken beschreven door dit model : de evolutie van levende systemen "convergeert" naar dit model. Het is het is als het ware de "grootste gemeenschappelijke beschrijving" van levende systemen, met andere woorden de synthese van de "evolutionaire gewoontes" van het leven.
De termen die gebruikt zijn voor het aanduiden van de verschillende subsystemen zijn vrij abstract, doch dit is bewust gedaan omdat ze moeten kunnen gelden voor elk potentieel levend systeem. Het onderzoek naar dergelijke algemene beschrijvingen van levende systemen, is gebeurd door interdisciplinaire werkgroepen waarin biologen, psychologen, biochemici, genetici, sociologen, economisten en ingenieurs samenwerken om een gemeenschappelijke visie te verkrijgen die over de verschillende disciplines heen begrijpelijk en aanvaardbaar is. In 1978 heeft James Grier Miller het resultaat van jarenlang interdisciplinair werk naar de definitie van levende systemen gebundeld in zijn voortreffelijke standaardwerk "Living systems".
De aanwezigheid van een Gamma-replicator hebben we al opgesomd als één van de meest typische eigenschappen.
We weten bijvoorbeeld ook dat het leven een "ver-van-evenwichtsstoestand" is. Om deze toestand te kunnen behouden, zal elk levend systeem zich moeten "voeden" met coherente materie/energie. Dit betekent dat hij een geschikte input moet bezitten om deze materie en energiebron binnen te brengen. Bovendien moet er een begrenzing zijn rond het levend systeem, die alle componenten van het systeem bij elkaar houdt. Deze begrenzing kan het systeem ook beschermen tegen vijandige druk van de omgeving, en kan selectief bepaalde inputs of outputs toelaten of beperken.
De voeding die is opgenomen, moet door verwerkt worden door een M/E-metabolisme, waarbij de energie wordt verbruikt voor eventuele voortbeweging of voor het intern metabolisme. De materiële voeding moet afgebroken worden en omgezet in structuren voor de opbouw van de verschillende "lichaamsdelen" van het levend systeem.
Om dit metabolisme te ondersteunen, moet een netwerk van kanalen bestaan om de materie en energie naar de juiste plaats in het systeem te brengen. Op de verschillende plaatsen in het systeem moet het dan verwerkt of geconverteerd worden in M/E-vormen die meer geschikt zijn voor de processen die plaatsgrijpen in de specifieke component van het systeem. In sommige gevallen zal er een overvloed aan voeding aanwezig zijn, waardoor het systeem over een opslagruimte moet bezitten, om de overtollige M/E te kunnen opslaan voor gebruik in periodes van schaarste. Dit metabolisme moet ervoor zorgen dat de geconverteerde - en al dan niet in de voorraad opgeslane- M/E wordt omgezet in stabiele componenten van het systeem. Zo moet de groei van het organisme worden ondersteund ; bepaalde componenten moeten op regelmatige tijdstippen worden vervangen of vernieuwd omdat ze aan slijtage onderhevig zijn, enz... Het neveneffect van dit metabolisme zal zijn dat de "weinig verspreide" M/E input geleidelijk aan wordt omgezet in enerzijds warmte, en anderzijds in een output van M/E die zodanig "verspreid" is, dat deze niet meer bruikbaar is voor het systeem. Dit betekent dat er aan het einde van de metabolische keten een subsysteem moet bestaan dat geschikt is voor de uitstoot (of extrusie) van deze niet meer bruikbare materialen. Dit ganse M/E metabolisme is dus eigenlijk een gesofisticeerde machine die entropie produceert, door omzetting van minder verspreide in meer verspreide M/E.
Deze ganse "metabolische machine" zal wellicht zo complex zijn, dat er speciale ondersteunende (supporterende) structuren nodig zijn om de ruimtelijke relaties tussen de subsystemen te garanderen. Zoniet zouden deze systemen kunnen instorten onder hun eigen gewicht. Deze ondersteuning zal moeten afgestemd zijn op de omgeving waarin het systeem leeft (water, lucht, land,...).
Sommige levende systemen zullen zo gesofisticeerd zijn, dat ze zich enkel met welbepaalde M/E bronnen willen of kunnen voeden. Dit betekent dat ze over een voortbewegingsmechanisme (een "motor") moeten kunnen beschikken, waarmee ze zich naar de voedingsbronnen kunnen begeven. Dit voortbewegingsmechanisme zal het systeem ook toelaten zich eventueel uit een vijandige omgeving te verwijderen, of bijvoorbeeld bevriende levende systemen op te zoeken. Ook de aard van dit voortbewegingsmechanisme zal sterk beïnvloed zijn door de omgeving waarin het systeem leeft.
Om een dergelijke complexe M/E -stroom te kunnen onderhouden, en te kunnen vrijwaren van natuurlijke en blinde verspreiding die alomtegenwoordig is, zal een parallelle informatiestroom moeten onderhouden worden die de complexiteit van de M/E stroom evenaart. Zoniet zou de complexe M/E stroom in de kortste keren gedoemd zijn tot een snel verval naar complete chaos, en de kracht van het specifieke metabolisme is net dat het verval naar de chaos sterk gemanipuleerd wordt, en daardoor vertraagd. Voor de ontwikkeling en replicatie van het systeem, is de informatie vervat als gecondenseerde Gamma-informatie. De ondersteuning van de overige functies van het systeem, die nodig zijn voor het normale levensonderhoud en levensbehoud, zullen diverse vormen van Alfa- en Beta, en in sommige gevallen ook Gamma- informatie worden benut.
Het zoeken naar de geschikte M/E bronnen impliceert bijvoorbeeld dat het levend systeem de voedselbronnen kan detecteren. Het moet dus over een sensorisch systeem ("input transducer") bezitten, dat toelaat de omgeving af te scannen naar mogelijk interessante plaatsen. Dit betekent dat informatie uit de omgeving moet kunnen binnengebracht worden in het levend systeem. Na het binnenbrengen in het levend systeem, moet deze informatie verwerkt worden op een wijze die analoog is aan de verwerking van de M/E stroom. Er bestaat dus ook een zekere vorm van "informatiemetabolisme".
Een eerste stap in dit informatiemetabolisme is de omvorming van de informatie die is binnengebracht, naar een M/E vorm die transporteerbaar is binnen het systeem. Daarna moet de informatie via de geschikte informatiekanalen, of eventueel via een informatienetwerk verspreid worden naar de relevante plaatsen binnen het systeem, zodat de informatie terecht komt, waar ze uiteindelijk kan worden verwerkt of gebruikt. Kenmerkend voor informatie is dat ze onder gecodeerde vorm kan worden overgebracht : Dit kunnen Alfa-, Beta- of Gamma- codes zijn. In dit geval moet de informatie vertaald (gedecodeerd) worden tot informatie die gebruikt kan worden waar ze nodig is.
Belangrijk voor meer complexe levende systemen is dat ze een leerproces kunnen doormaken, waarbij gedurende hun levenscyclus nuttige ervaringen en kennis uit het verleden, kunnen opslaan om in de toekomst te gebruiken. Om dit te kunnen, moet een subsysteem bestaan dat verbanden of associaties kan leggen : het systeem moet bijvoorbeeld een verband leggen tussen een bepaalde plaats, en een bepaald kenmerk, bijvoorbeeld dat ze rijk is aan voedselbronnen. Net als de M/E moet kunnen worden opgeslaan voor een eventuele latere behoefte, moeten de informatie en de associaties worden opgeslagen voor later gebruik : dit gebeurt in het geheugen. Voor de besturing van het ganse systeem moeten alle relevant inputs en associaties, al dan niet afkomstig uit het geheugen, gecombineerd worden tot een beslissing of output die de handelingen van het systeem controleert. Op basis van kennis opgeslagen uit het verleden, kan het systeem beslissen om de huidige verblijfplaats te verlaten, en een plaats op te zoeken die in het geheugen is gegrift als zijnde rijker aan voeding. Dit ganse gebeuren (beslissing en coördinatie van de verplaatsing) gebeurt door het beslissingscentrum (de "decider"). Zonder dit subsysteem zou coördinatie van het levend systeem onmogelijk zijn, en zou een snel verval tot chaos niet te vermijden zijn.
In sommige gevallen moet de informatie uit het beslissingscentrum "gecommuniceerd" worden naar de omgeving van het systeem. Dit betekent dat de informatie eventueel opnieuw moet gecodeerd worden, naar een vorm die verstaan wordt door de omgeving. Deze gecodeerde vorm moet daarna overgebracht worden naar de omgeving, door een communicatiesysteem dat voor de output zorgt (de "output transducer").
Eens de output geproduceerd is, heeft de informatiestroom het levend systeem opnieuw verlaten. Deze informatiestroom kan hierbij in complexiteit zijn toegenomen. Dit gaat echter ten koste van energieverbruik van de M/E stroom, waardoor het netto-effect toch een toename van de "verspreiding" van de M/E en I-stroom zal zijn.
Zoals eerder gesteld zijn levende systemen "open systemen", die voortdurend te maken hebben met variaties in de flux van materie, energie en informatie. Ze trachten tussen al de tegenstrijdige variabelen in hun omgeving, een compromis te vinden en zo een "quasi-evenwicht" of "homeostase" te bereiken. Dit quasi-evenwicht trachten ze te behouden door een goede balans tussen de subsystemen die materie, energie en informatie verwerken, en dit in harmonie met de systemen van een hoger niveau in de hiërarchie, en aangepast aan de omgeving. Hoewel er voor elke "proces-variabele" een range van stabiliteit is, zullen bepaalde variaties van de stroming van materie, energie of informatie toch een zekere stress binnen het levend systeem veroorzaken.
Als complexe "chemische fabriek" zal het levend systeem op deze stress reageren volgens het "principe van Le Chatelier" : "Elk systeem in chemisch evenwicht zal bij een variatie van één van de evenwichtsfactoren, zodanig reageren dat het een variatie doet ontstaan die tegengesteld is aan de factor in kwestie". Dit wordt veelal breder geformuleerd als "Een stabiel systeem onder stress zal in een dergelijke richting bewegen, dat de stress wordt geminimaliseerd". Dit klinkt ondertussen bekend in de oren als één van de basiskenmerken van het principe van de fluxmaximalisatie.
Levende systemen zijn specialisten in het minimaliseren van de gevolgen van stresscondities : het zijn de gradiëntdissipatoren bij uitstek.Ze bezitten tal van regelingsprocessen die de belangrijkste variabelen moeten stabiliseren, ondanks de stresscondities. Deze feedbackcycli en hypercycli zijn zodanig met elkaar verweven, dat een levend systeem een verbazende stabiliteit vertoont, ondanks (of dankzij) zijn hoge complexiteit en zijn situatie ver uit evenwicht. Zo wordt bijvoorbeeld de totale hoeveelheid vaste stoffen en vloeistoffen binnen een levend organisme, op een constant niveau gehouden, met een zekere oscillatie rond een gemiddelde waarde. Dit is maar één van de talrijke functies van een organisme, waarbij een toestand van quasi-evenwicht wordt behouden door het gebruik van externe energie, die uiteindelijk afgeleid is van zonne-energie.
Ik zal deze inleiding over de levende systemen beëindigen met de boutade "waar voeding is, zal een levend systeem ontstaan". Met deze "mindcatcher" wil ik erop wijzen dat het niet alleen waarschijnlijk is dat de levende systemen in de evolutie ontstaan ; het is ook waarschijnlijk dat ze hierbij de verschillende de ecologische niches zullen vullen. Dit is bovendien in overeenstemming met het principe van de fluxmaximalisatie. De aanwezigheid van voeding impliceert immers een energie-gradiënt, die door één of andere vorm van gradiënt-dissipatie zal worden tenietgedaan.
Terug naar de index van dit hoofdstuk
3.2. Enkele voorbeelden van levende systemen
Net als we bij elementaire deeltjes verschillende niveau’s kunnen herkennen, waarbij een bovenliggend niveau is samengesteld uit componenenten van een onderliggend niveau (bijvoorbeeld subatomaire deeltjes, atomen, molecules, ....planeten, sterrenstelsels, clusters van sterrenstelsels ...), kunnen we bij levende systemen dergelijke niveau’s onderscheiden. Naargelang de criteria die men hanteert, kan men meer of minder niveau’s onderscheiden.
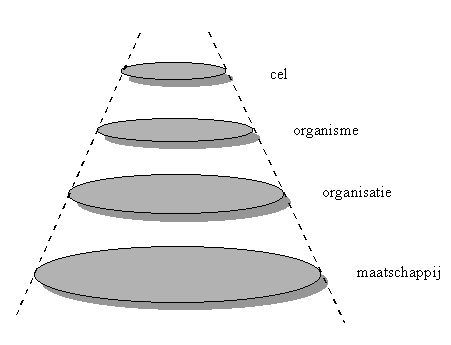
Figuur H7F9 : enkele voorbeelden van levende systemen, die beantwoorden aan de kenmerken van de Universele Gamma-attractor
Bij de levende systemen op aarde kunnen we bijvoorbeeld cellen onderscheiden. Deze cellen vormen samen levende organismen zoals planten, dieren en mensen. De mensen kunnen zich groeperen tot menselijke organisaties vormen, die op hun beurt gebundeld kunnen worden tot maatschappijen. Elk van deze vier niveau’s heeft de kenmerken van levende systemen. Dit zal in de volgende paragrafen worden verduidelijkt.
We zullen hier kort enkele kenmerken van de M/E stroom en de I stroom toelichten, om aan te tonen hoe subtiel de stromen van materie,energie en informatie geworden zijn, in vergelijking met bijvoorbeeld de Beta-systemen.
Doelstelling hierbij is ook aan te tonen dat een levende cel, een organisme, een organisatie en een maatschappij inderdaad de verschillende subsystemen en processen bevat, die in de algemene beschrijving van de Universele Gamma-Attractor (kortweg "UGA") zijn vermeld.
Terug naar de index van dit hoofdstuk
Figuur H7F10 : enkele voorbeelden van cellen
Biologische cellen bestaan er in vele maten en vormen. Sommige cellen kunnen volledig op zichzelf bestaan, en worden micro-organismen genoemd. Hieronder bevinden zich micro-organismen die infectueuze ziekten kunnen veroorzaken bij plant, mens en dier, doch er zijn ook micro-organismen die een positieve invloed hebben op de mens, zoals bij het maken van bier, wijn en andere gefermenteerde producten. Andere cellen maken deel uit van hogere organismen, zoals witte en rode bloedcellen of cellen in planten. Deze zijn niet volledig zelfstandig, en kunnen enkel bestaan als een deel van het grotere organisme waartoe ze behoren. We zullen ons voornamelijk richten op de vrijlevende cellen.
De Gamma-replicator wordt gevormd door het DNA, samen met een aantal ondersteunende organellen (de "organen van de cel") zoals de ribosomen, mitochondria,... De cruciale gebeurtenis in de cellulaire reproductie is de vertaling van de genetische informatie uit het DNA, naar de de volgende generatie. De cellulaire replicatie kan zowel sexueel als a-sexueel zijn. Bij a-sexuele replicatie, zoals die gebeurt bij bepaalde bacteriën, protozoa, wordt het DNA in de cel gecopieerd en een kleine, minder gedifferentieerde cel wordt afgescheiden. Er wordt dus een kloon gevormd, waarbij de genetische informatie identiek blijft aan deze van de voorouder. Bij de sexuele reproductie komen 2 cellen dicht bij elkaar, waarbij hun celmembranen nauw met elkaar in contact zullen komen (op een manier die gelijkaardig is met de manier waarop cellen in weefsels met elkaar binden). Hierbij wordt de helft van de genetische informatie van de ene cel gecombineerd met de helft van de genetische informatie van de andere cel. Voordeel van de sexuele reproductie is dat de reproductie meer variatie brengt in de genetische informatie, waardoor de natuurlijke selectie bevorderd wordt. De kans bestaat immers dat de nieuwe cel eigenschappen heeft, die ze meer aangepast maakt aan de omgeving, waardoor de overlevingskansen groter zijn.
De begrenzing van de cel wordt gevormd door een membraan, die zowel een begrenzing vormt voor de M/E stroom, als voor de I-stroom in en uit de cel. Door de specifieke samenstelling is de membraan selectief doorlaatbaar : hierdoor laat ze specifiek bepaalde M/E componenten door, en verhindert ze dat ongewenste componenten de cel binnen of buiten komen.
De informatietransfer doorheen de membraan wordt beïnvloed door de membraanpermeabiliteit en het concentratieverschil tussen componenenten binnen en buiten de cel,op dezelfde wijze als dit geldt voor de ME/-transfer, althans als het Alfa-informatie in chemische molecules betreft. De concentratieverschillen tussen molecules (zoals Natrium- of Kaliumionen) veroorzaakt een reguleringsproces dat gebaseerd is op Beta-informatie. Overmatige concentratieverschillen vormen een signaal van Beta-informatie, die een flux van ionen in gang zet, die het concentratieverschil zal verminderen tot het aanvaardbaar klein is (een mooi voorbeeld van gradiëntdissipatie).
- M/E stroom :
Wat betreft het M/E metabolisme, moet in een eerste stap de juiste M/E worden binnengebracht. Vrijlevende cellen moeten op zoek gaan naar voedsel. Bij amoebes bijvoorbeeld zal de celmembraan, wanneer deze in conctact komt met voedsel, deze het voedsel langzaam omcirkelen met zijn pseudopodia. De pseudopodia (de ingestor) worden gevormd door uitbreidingen van de celmembraan en het cytoplasma (celvocht). Het voedsel wordt volledig ingesloten door celmembraan, en wordt opgenomen in de cel. Binnen deze membraan kunnen de metabolische processen reeds starten. Andere vrijlevende cellen bezitten over andere mechanismen voor opname van voedsel. Opgeloste materialen kunnen de cel binnenkomen door een analoog proces : dit "drinkproces" wordt "pinocitose" genoemd. Water daarentegen kan op alle plaatsen door de celmembraan diffunderen. Opgeloste gassen, zoals zuurstof voor de ademhaling, dringen door de membraan zoals bij het pinocitose-proces.
Van zodra de voedingsstoffen zijn opgenomen, moeten ze verspreid worden over de cel. Binnen vele cellen gebeurt dit door een basisstroom van het cytoplasma binnen de cel : al het celvocht stroomt in één welbepaalde richting, of in 2 tegengestelde richtingen. Het endoplasmatisch reticulum is een voorbeeld van een organel gespecialiseerd in de distributie (een distributor) van metabolische stoffen.
Voor de verwerking van de voedingsstoffen zijn de mitochondria de gespecialiseerde organellen (de convertor). Het zijn staafvormige structuren, omgeven door een dubbele membraan, waarbij de inwendige membraan inwendig is opgevouwen als een lasagne. Binnenin het mitochondrion, zijn de enzymes (de biochemische machines) op specifieke wijze geordend als een soort moleculaire assemblagelijn. Hun plaatsing komt overeen met de volgorde van de chemische reacties die ze moeten uitvoeren en controleren. Deze die energierijke molecules (zoals ATP, de universele brandstof van de cel) moeten aanmaken, zijn gelegen aan de binnenkant van het mitochondrion, terwijl deze die energieverslindende syntheses moeten uitvoeren, bij de buitenkant van het membraan zijn gelegen. Een cel kan in totaal 6 tot 5000 mitchondria bezitten, die gelegen zijn op de plaatsen waar de conversieproducten het meest benodigd zijn. De mitochondria zijn te beschouwen als de ademhalingsapparaten van de cel : ze oxyderen de voedingsstoffen tot een brandstof die binnen de ganse cel kan gebruikt worden voor chemische reacties in alle delen van de cel, zoals de vorming van eiwitten. De omzetting in ATP gebeurt voornamelijk om de energie-overdracht bij de verdere chemische reacties efficiënter te laten verlopen. Op deze wijze wordt een energieconversie bereikt met een efficiëntie tot 66% : wetende dat de efficiëntie van energiecentrales op basis van stoomproductie ongeveer 30% is, is dit verbazend hoog. Bij deze conversieprocessen kunnen complexe biochemische cycli een belangrijke rol spelen, zoals de glycolyse en de Krebbscyclus of citroenzuurcyclus. Beiden zijn zeer mooie voorbeelden van zelfregulerende processen of processen met Beta-attractoren (cfr. vorig hoofdstuk in verband met biochemische cycli). Het gehele energieconversieproces is een schitterend voorbeeld van koppelingen, terugkoppelingen, kanalisaties en de vorming van hypercycli in levende systemen. We vinden er dus de diverse niveau’s van cybernetische Beta-systemen in terug.
Wanneer de basisproducten uit de voedingsstoffen zijn gevormd, kunnen ze verder verwerkt worden in producten nodig voor de groei of herstelling van beschadigde componenten. Dit is het "productie-"stadium van het metabolisme. Alle cellen produceren eiwitten, koolhydraten, energierijke fosfaten, vetten en andere bestanddelen. Deze productie wordt uitgevoerd en gecontroleerd door enzymes. Een cruciaal productieproces voor alle levende systemen op aarde is bijvoorbeeld de fotosynthese, die wordt uitgevoerd door sommige ééncelligen, doch ook door chlorofylhoudende (groene) planten. Hierbij wordt de energie van het zonnelicht geconverteerd in energierijke verbinding zoals ATP, en wordt zuurstof afgegeven aan de omgeving. Dit proces is cruciaal voor de evolutie van levende wezens om twee redenen :
- het zonnelicht is wellicht de belangrijkste energiebron voor alle leven op aarde, doch de meeste levende systemen kunnen zonne-energie niet rechtstreeks als energiebron gebruiken ; ze moeten dit zonnelicht dus indirect consumeren door voeding met fotosynthetische levende systemen,
- door het onstaan van de fotosynthese, is de aardatmosfeer geleidelijk aangerijkt met zuurstof ; als gevolg hiervan zijn geleidelijk aan zuurstofverbruikende organismen (waaronder de mens) ontstaan..
De productie van eiwitten, koolhydraten en andere componenten vergt energie. Om deze reden moet de cel voor periodes van schaarste een energievoorraad opslaan, alsook een voorraad aan andere bouwmaterialen. Energie wordt meestal opgeslagen onder twee vormen, een snel toegankelijke en een traag toegankelijke vorm. De snelle vorm is de reeds vermelde ATP ; tragere vormen van opslag vinden we terug in vetten en koolhydraten. Deze verschillende types voorraden laten toe de energieconversiesnelheid in de cel te laten variëren volgens de noodzaak.
Na de conversie- en productiefase, zullen bepaalde producten uit de cel moeten verwijderd worden. Het verkeer door de celmembraan is dus een twee-weg proces. De hoeveelheid water bijvoorbeeld wordt in evenwicht gehouden doordat het door beide zijden van de membraan kan diffunderen, als respons op de concentratiegradiënt. Ook sommige electrolyten en andere bestanddelen zoals koolstofdioxide en zuurstof diffunderen (in opgeloste toestand) doorheen beide zijden van de membraan. De extrusiefunctie fungeert als het inverse van de ingestorfunctie. Veelal zijn beide processen aan elkaar gekoppeld. Een Kalium-ion kan een cel enkel binnentreden, als ze tegelijkertijd wordt uitgewisseld tegen een Natrium-ion. Sommige hogere ééncelligen beschikken reeds over meer gespecialiseerde structuren voor de extrusie van bepaalde stoffen naar de omgeving. Een agressief voorbeeld vinden we bij de netelcellen : deze zijn in staat, bij prikkeling door een vermeende vijand, een ganse zweepdraad naar buiten toe te laten exploderen, met de bedoeling de vijand schade toe te brengen.
Dit ganse M/E metabolisme veronderstelt dat voldoende voedingsstoffen worden binnengebracht in de cel. In sommige situaties zal de cel zich moeten voortbewegen om bij de voedingsstoffen te komen.
Een mooi voorbeeld van een motorische beweging gedreven door het cytoplasma vinden we terug bij de amoebe. Wanneer een amoebe beweegt, vloeit het plasma in de bewegingsrichting. Aan het uiteinde van de beweging vloeit het langs de wanden van de cel, en verandert in een meer vaste, moeilijker vloeibare structuur. Aan de andere kant gebeurt de omgekeerde transformatie. Terwijl dit gebeurt, wijzigt de vorm van de amoebe, en worden pseudopodia (schijnvoeten) gevormd in de richting waarin wordt bewogen. Deze amoeboïdale beweging is ook van belang bij hogere organismen. Gedurende de embryonale ontwikkeling van hogere organismen, bewegen cellen - die later in een weefsel zullen gefixeerd worden - naar hun uiteindelijke positie via een amoeboïdale beweging. Gedurende de verdere differentiatie van de cel, verliest ze de capabiliteit om te bewegen.
Andere ééncelligen bewegen door middel van zweepdraden, zoals de zaadcellen bij de mens dit doen. Bij hogere organismen bestaan cellen die gespecialiseerd zijn in de voortbeweging, zoals bijvoorbeeld de spiercellen.
Al deze kenmerken van de M/E stroom zouden onmiddellijk vervallen wanneer de cel zijn ruimtelijke structuur niet stevig ondersteunt.
Studies van de levende cel tonen aan dat er een vorm van "cytoskeleton" bestaat, die fungeert als de supporterstructuur. Dit "skelet" wordt opgebouwd uit een netwerk van kleine buisjes en kanaaltjes (de "microtubuli en microfilamenten") en maakt bovendien deel uit van de distributor en de motor.
- I- stroom :
De ganse M/E stroom moet natuurlijk gecoördineerd worden door een complexe informatiestroom. Sommige cellen, naast de vrij-levende cellen, zijn zelfs volledig gespecialiseerd in informatieverwerking. Tot deze klasse behoren bijvoorbeeld de neuronen (in de hersenen van hogere dieren) en de receptoren.
Net zoals de eerste stap in het M/E metabolisme het binnenbrengen van de M/E is, is de eerste stap in de informatieverwerking het binnenbrengen van de informatiedragers via de input transducer. De meeste vrijlevende cellen hebben hier geen gespecialiseerde structuren voor, en de informatiedragers worden op analoge wijze als de M/E voedingsstoffen binnengebracht. Enkele Flagellaten zoals Euglena beschikken over receptoren die lichtgevoelige pigmenten bevatten, gelijkaardig aan deze in receptorcellen van de ogen van hogere organismen. Zoals reeds gezegd bevatten hogere organismen wel tal van gespecialiseerde receptorcellen, zoals drukreceptoren, chemoreceptoren, geurreceptoren smaakreceptoren, lichtgevoelige receptoren, enz... Deze receptoren werken veelal op het "sleutel op het slot"- principe dat kenmerkend is voor Alfa-informatie.
Van zodra deze informatie is binnengebracht in de cel, moet ze vertaald worden naar een M/E vorm die in de cel makkelijk kan worden getransporteerd. Eiwitten zijn bijzonder geschikt voor deze interne informatieoverdracht. Bij het binden met de informatiedrager, kan hun ruimtelijke structuur van vorm veranderen, waarbij deze vormverandering geldt dan als interne informatieoverdracht. Bacteriën kunnen zo bijvoorbeeld de productie van enzymes afstemmen op de concentratie van voedingsstoffen in het milieu. In afwezigheid van een bepaalde voedingsstof (het substraat) wordt het enzym dat instaat voor verwerking ervan niet aangemaakt. Is het substraat wel aanwezig, dan wordt het enzym in grote hoeveelheden geproduceerd.
Van zodra dat deze informatie vertaald is naar een transporteerbare vorm, kan ze gedistribueerd worden binnen de cel. De molecules die drager zijn van Alfa-gecodeerde informatie worden op dezelfde wijze getransporteerd als de andere M/E componenten. Sommige cellen bezitten speciale kanalen voor overdracht van Beta-informatie. De algen Nitella zijn hierdoor in staat electrische stromen te geleiden. Neuronen (hersen- en zenuwcellen) zijn zodanig gespecialiseerd in het transport van electrische inpulsen doorheen hogere organismen, dat ze als lange dunne kabels doorheen het lichaam van deze organisme kunnen liggen. Hun celmembraan is zodanig gedifferentieerd, dat electrische impulsen optimaal worden overgedragen.
Wanneer deze informatie is overgedragen naar de finale bestemming, moet ze eventueel gedecodeerd worden in een "locale" code. Alfa-informatie is meestal bepaald door de chemische structuur van de drager-molecule. Deze kunnen bijvoorbeeld op specifieke moleculaire bindingsplaatsen gaan binden. Deze bindingsplaatsen herkennen de chemische structuur, en bij de binding wordt een electrisch signaal gevormd, of wordt een chemische reactie geïnitieerd. Beta-informatie kan getransporteerd zijn onder de vorm van lichtsignalen, temperatuursgradiënten, drukken, enz... Deze signalen kunnen op hun beurt aanleiding geven tot een ander Beta-signaal.
Een nadeel van de verschillende hercoderingen van de informatie, is dat een deel van de informatie verloren gaat door de onherroeppelijke verspreiding, die ook kenmerkend is voor de informatiestroom.
Of vrijlevende cellen deze informatie gebruiken voor een leerproces, zoals hogere organismen dit kennen, staat nog onder discussie. Experimenten met hogere ééncelligen (Paramecia) tonen aan dat ze in staat blijken te zijn voedselrijke zones in hun milieu te herinneren, na eruit verwijderd te zijn. Het staat echter ter discussie of dit moet beschouwd worden als een resultaat van snelle en effectieve ad hoc informatiedoorstroming, dan wel als het resultaat van een primitief leerproces. Elk leerproces vereist ook de aanwezigheid van een geheugen. De Gamma-replicator, met zijn (genetische) informatie, is een duidelijke vorm van een geheugen. Verschillende andere functies in de cellen zijn te vergelijken met de werking van een geheugen, in die zin dat een informatie-input een langdurige wijziging veroorzaakt van bepaalde informatiedragers. Een voorbeeld dat hierboven reeds werd aangehaald is de enzyme-inductie door aanwezigheid van specifieke substraten. Bij aanwezigheid van specifieke substraten, zullen bepaalde micro-organismen de instructies opvragen uit hun geheugen, om de enzymes te maken om dit substraat af te breken. Dit betekent dat een "bibliotheek" van instructies bestaat, die geraadpleegd wordt wanneer daar behoefte aan bestaat. Dit betekent dat de betrokken enzymes enkel worden aangemaakt wannneer ze nodig zijn, wat een aanzienlijke kostenreductie impliceert. Deze uiting van het "principe van de minimale inspanning" of van het "economisch principe" is een duidelijk competitief voordeel bij de natuurlijke selectie van deze organismen.
In deze overvloed aan informatie, moeten regelmatig beslissingen worden genomen voor verdere coördinatie van de activiteiten van de cel. Het beslissingscentrum (de "decider") van de cel bestaat volgens Eigen uit 2 niveau’s, die hij de wetgevende macht en de uitvoerende macht noemt. De wetgevende macht bestaat uit de nucleïnezuren in het genetisch materiaal. De genen bevatten alle programma’s of wetten die de cellulaire organisatie bewaken. Deze wetten moeten vertaald worden naar een uitvoerend niveau, het niveau van de enzymes die specifiek chemische reacties controleren en uitvoeren. Een experiment op vlak van "beslissen in onzekerheid" kan uitgevoerd worden met bacteriën. Zo werd de beweeglijke bacterie E. Coli geplaatst in een milieu met aan de ene kant een aantrekkelijke voedingsstof en aan de andere zijde een afstotende stof. De bacterie bevindt zich in een gradiënt van deze stoffen, en moet beslissen welke kant ze uitgaat. De bacterie krijgt tegenstrijdige signalen vanuit zijn chemoreceptoren, vergelijkt deze signalen met zijn geprogrammeerde regels voor overleving. Ze moet dan een beslissing treffen en een commando geven aan het voortbewegingsmechanisme (de flagella) om te bewegen in de richting van de voedingsrijke kant. Bij een dergelijke beslissing is een hele reeks reacties tussen chemische componenten betrokken. Levende cellen zijn zo complex omdat ze een heel gamma van dergelijke reactie- en controlemechanismen hebben opgebouwd gedurende hun cellulaire evolutie.
Van zodra een beslissing is genomen, moet deze soms nog gecodeerd word tot een vorm die verstaanbaar is op de plaats waar de beslissing wordt uitgevoerd. Een bijzonder voorbeeld hiervan komt voor bij het ééncellig organisme Plasmodium. Deze scheidt een stof af (acrasine) die ontvangen wordt door andere Plasmodium cellen. Dit is het signaal voor aggregatie van de cellen tot een meercellige massa. Door dit mechanisme bevindt de cel zich op de grens van ééncellig en meercellig organisme.
Dit mechanisme start in stress-omstandigheden, zoals gebrek aan voeding, en de massa vormt zich om tot een plantachtig aggregaat, met een stengel en vertakkingen, die op hun uiteinden vruchtlichamen kunnen dragen. Dit is één van de mooie voorbeelden van zelforganisatie en stress-demping, waarbij voorbij een kritisch punt (een te grote honger), de hypnonen of "slaapwandelende" cellen "wakker" worden, en na een vorm van vergaande commincatie zich organiseren over de eenheden door, zonder dat dit door een centraal brein wordt gecoördineerd. Deze nieuwe organisatie is beter aangepast aan de stress-condities van de omgeving, en biedt betere overlevingskansen.
Dit is ook een voorbeeld waar de Alfa-informatie finaal buiten de cel moet getransporteerd worden.
Deze Plasmodium-beschrijving, die naargelang de situatie ééncellig of meercellig is, is ook een geschikt ogenblik om over te stappen naar een hoger niveau in de hiërarchie van de levende systemen, dit van de multicellulaire organismen.