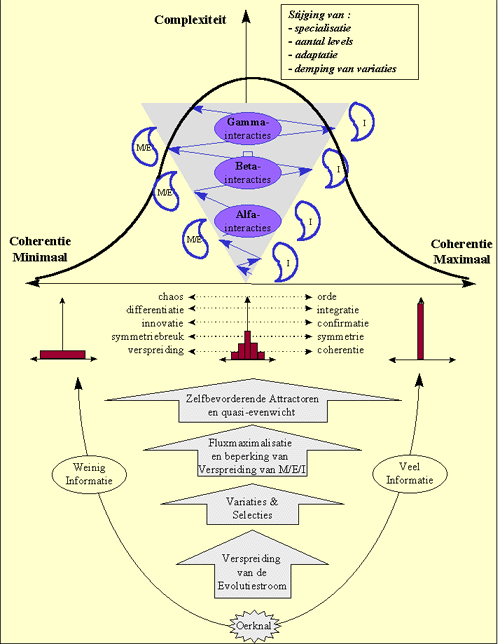
In de vorige hoofdstukken werd uitvoerig aangetoond dat de evolutie gekenmerkt is door een kwantitatieve en kwalitatieve verspreiding van materie, energie en informatie.
Bij deze verspreiding worden bepaalde toestanden geselecteerd en andere niet, waardoor de evolutie kan aanzien worden als een zich continu vertakkend netwerk van evolutiewegen. Naarmate we hierbij verder gaan in de evolutie, neemt de symmetrie van de evolutiepatronen geleidelijk aan af. Dit gaat zover dat we uiteindelijk slechts nog maar een "gemiddelde symmetrie" kunnen waarnemen, bij het herhalen van evoluties.
Hierbij werd uitvoerig ingegaan op het belang van informatie in de evolutie.
In afwezigheid van informatie, wordt de meest waarschijnlijke toestand geselecteerd. Dit nemen we bijvoorbeeld waar bij een druppel inkt, die zich in een glas water verspreidt : de blinde verspreiding leidt tot een toestand met maximale verspreiding, precies omdat dit de meest waarschijnlijke toestand is. De evolutie gaat hierbij in één welbepaalde richting, namelijk deze van de grootste waarschijnlijkheid.
Anders wordt het echter wanneer informatie in het systeem aanwezig is.
De aanwezigheid van informatie beïnvloedt de selectie in de evolutie.
Naarmate de hoeveelheid informatie stijgt, neemt de "blinde" verspreiding in de evolutie af. De aanwezige informatie verhoogt de waarschijnlijkheid van bepaalde evolutiewegen, en verlaagt de waarschijnlijkheid van andere. De informatie trekt de evolutie in een bepaalde richting : ze fungeert als een "attractor" (aantrekker) voor bepaalde evolutiepatronen (zoals een rode vlag een stier aantrekt). Naarmate er meer informatie aanwezig is, wordt dit effect meer uitgesproken. Niet alleen de hoeveelheid informatie, doch ook het type informatie is doorslaggevend voor de organisatie van de evolutiepatronen.
Als deze informatie een selecterende betekenis heeft in het ganse universum, dan kunnen we spreken van een "universele attractor". Hiertoe behoren de 4 basiskrachten, en ook bijvoorbeeld de universele Gamma-attractor.
Als deze informatie slechts een betekenis heeft binnen een beperkte context van het universum, hebben we dit een "contextuele attractor" genoemd. Naarmate de informatie bepaald wordt door de materie, door een proces in de tijd, of door symbolische relaties, hebben we dit Alfa-, Beta- of Gamma-attractoren genoemd. Deze attractoren leiden elk tot interactoren met duidelijk verschillende kenmerken.
In dit hoofdstuk zullen we dieper ingaan op de diverse types selectie die er kunnen onderscheiden worden in de evolutie. De selectie is immers niet enkel afhankelijk van het type informatie dat aanwezig is, doch ook nog van een aantal andere factoren.
In een eerste paragraaf zullen we deze factoren eens op een rijtje zetten, om daarna meer verduidelijking te geven met een aantal concrete voorbeelden.
Terug naar de index van dit hoofdstuk
1. Variatie en selectie in de evolutie
Het principe van de variatie, opgevolgd door de selectie, is een basisprincipe in de evolutie.
Ook dit basisprincipe zelf is echter ge-evolueerd. De manier waarop bijvoorbeeld een auto onstaan is, is sterk verschillend van de wijze waarop een giraf is ge-evolueerd. Dit is op zijn beurt niet te vergelijken met de blinde verspreiding van de gasatomen in een gas. Nochtans maken al deze voorbeelden deel uit van de evolutie van het universum, en moet het dus mogelijk zijn om naast de verschilpunten in evolutieprincipes, ook gelijkenissen aan te treffen. Dit zullen we in de volgende paragrafen doen voor wat betreft de principes van variatie en selectie.
Opnieuw speelt informatie een sleutelrol. Zowel het gas, als de giraf en de auto zijn ontstaan door een proces van variatie en selectie ; het verschil zit in het al dan niet aanwenden van voorkennis of informatie bij de variatie en de selectie.
Dit is verduidelijkt in onderstaande grafiek.
Figuur H8F1 : in de loop van de evolutie zijn verschillende evolutieprincipes onstaan
Het is duidelijk dat in de vroege evolutie, er geen of nauwelijks informatie aanwezig was. Variaties in het gedrag van evolutiepatronen waren enkel en alleen te wijten aan de blinde verspreiding. Ook was de selectie van evolutiepatronen volledig blind, wat resulteerde in de selectie van de meest waarschijnlijke evolutiepatronen. Dit komt overeen met het "selectieprincipe 1" in de grafiek, dat ik de "blinde evolutie" heb genoemd.
Geleidelijk aan onstonden door dit proces van blinde variatie en selectie, interactoren met een zekere betekenis in het universum of binnen een bepaalde context. Deze eerste interactoren hadden een selecterende invloed op de evolutie, en versterkten duidelijk bepaalde evolutiewegen ten nadele van andere.
Bij het proces dat Darwin "natuurlijke selectie" heeft genoemd, vinden we echter zowel processen terug die gelijken op "blinde selectie" als van processen die zeer duidelijk behoren tot de "geïnformeerde selectie". Als een bepaalde diersoort uitsterft door een natuurramp, leunt dit nauw aan bij een blinde selectie : het onstaan van de diersoort is echter in vele stadia van de evolutie geïnformeerd. Om die reden zal ik het bij "geïnformeerde" selectie catalogeren. Ontstaat echter bijvoorbeeld een nieuwe diersoort door een proces van "sexuele selectie" en isolatie, dan is dit een duidelijk voorbeeld van geïnformeerde selectie : het is een gevolg van de gerichte (geïnformeerde) partnerkeuze die generaties na generaties consequent is doorgevoerd.
Bij een proces van zelforganisatie (zoals we bij de Beta-interactoren hebben besproken), onstaat informatie die nieuwe variaties mogelijk maakt, die voordien niet mogelijk waren. Het zijn als het ware emergente variaties, ontstaan door het proces van spontane zelforganisatie. De informatie is hier impliciet aanwezig gedurende het zelforganiserende proces, waardoor we kunnen spreken van geïnformeerde variaties. Deze geïnformeerde variaties, worden samen met de toevallige variaties, geselecteerd in een evolutieproces dat door Stuart Kaufmann "zelforganisatie en selectie" werd genoemd. In werkelijkheid lopen de natuurlijke selectie (het "Darwinistische proces") en de zelforganisatie en selectie (het "Kaufmannproces") interactief en nauwelijks te onderscheiden door elkaar : vandaar dat ik het gezamelijke proces "zelforganisatie en natuurlijke selectie" heb genoemd. Beide principes vullen elkaar dus aan en ageren complementair.
Op aarde is de mens in heel expliciet in staat tot het verrichten van "ge-informeerde" variaties. In een biotechnologisch labo kan men bijvoorbeeld aan het genetisch materiaal van een kolonie bacteriën, extra genetisch materiaal toevoegen dat de bacterie resistent maakt tegen bijvoorbeeld een antibioticum. Wanneer we deze kolonie bacteriën naderhand in contact brengen met een oplossing die het betrokken antibioticum bevat, zullen bepaalde bacteriën afsterven, en andere niet. Het variatieproces is hier geïnformeerd door de mens.
Het gaat echter verder dan een geïnformeerde variatie. Ook het selectieproces van het afsterven is gedeeltelijk door de mens bepaald, waardoor dit een voorbeeld is van "selectieprincipe 3", of de "artificiële constructie". Er zijn echter voorbeelden, waarbij de invloed van de informatie op een selectie die volgt op een geïnformeerde variatie, nog veel duidelijker is.
Wanneer een autoconstructeur een nieuwe auto ontwerpt, gebeuren de variaties op het "auto-thema" vanuit de voorkennis van wat de klant verwacht van een auto bij zijn aankoop. De variaties bij het ontwerp van de auto zijn dus geïnformeerd. De selectie van de finale autodesign die op de markt zal worden gebracht, gebeurt vanuit dezelfde voorkennis van de verwachtingen van de finale eigenaar van de wagen. Hetzelfde geldt voor de meeste artefacten die door de mens zijn geconstrueerd ; vandaar dat ik dit de "artificiële constructie" zal noemen.
In de volgende paragrafen zullen we dieper ingaan op elk van deze principes. We zullen hierbij tot het inzicht komen dat alles wat we waarnemen, is ontstaan door één van deze selectieprincipes.
Terug naar de index van dit hoofdstuk
2. Blinde evolutie in het universum
Wanneer variaties in complete afwezigheid van informatie ontstaan, is er geen enkele "voor"keur voor of "voor"kennis over een bepaalde evolutierichting. Op dit meest fundamentele niveau, "weten" informatieprocessen niet welke richting er zal geselecteerd worden.
Wanneer een blinde variatie ontstaat, zijn er steeds meerdere mogelijke variaties of uitkomsten van het variatieproces.
Of een variatie zal worden geselecteerd, hangt af van de "stabiliteit" van de eruit voortvloeiende interactoren : stabiele interactoren worden weerhouden (geselecteerd), en onstabiele worden ge-elimineerd .
Dit principe is zelfbevestigend, omdat "stabiliteit" net kan gedefinieerd worden als datgenene wat niet (snel) verdwijnt of verandert : het is dus bovendien een zeer relatief begrip, dat afhangt van de context waarin het wordt gebruikt.
Door gebruik te maken van het "gasmodel" werd in één van de vorige hoofdstukken reeds aangetoond dat deze stabiliteit bij niet-geïnformeerde systemen ook precies overeenkomt met de hoogste waarschijnlijkheid.
Bovendien is het zo dat de waarschijnlijkheid van een evolutie van een minder waarschijnlijke toestand naar een meer waarschijnlijke toestand, groter is dan de waarschijnlijkheid van het omgekeerde.
Deze heel eenvoudige bewering, is eveneens van fundamenteel belang voor de evolutie. Dit principe impliceert immers een fundamentele assymetrie in de evolutie.
Door dit principe zal de evolutie immers steeds in een welbepaalde richting gaan (gemiddeld genomen), en kan de richting van de tijd worden vastgelegd (zoals in vorige hoofdstukken reeds aangehaald).
De evolutie is dus gemiddeld gezien "assymetrisch", omdat bepaalde richtingen bevoordeeld worden.
Doordat bepaalde richtingen met een grotere waarschijnlijkheid worden gekozen, is dus de tijdssymmetrie gebroken : de evolutie gaat niet evenveel "vooruit" als "achterruit" : ze gaat voornamelijk vooruit. En, wat meer is, gemiddeld gezien kan de evolutie niet omgekeerd worden, precies ten gevolge van dit proces van blinde variatie en blinde selectie.
Blinde variatie en blinde selectie zijn dus de basis van de assymetrie in de evolutie.
Ze zijn hierdoor ook de oorzaak van de richting van de tijd, en van de gemiddelde onomkeerbaarheid in de gemiddelde evolutie.
Of hoe gebrek aan voorkennis en informatie, precies de oorzaak kunnen zijn van een schijnbaar door alwetendheid gedreven besturing van de evolutie.
Terug naar de index van dit hoofdstuk
3. Ge-informeerde selectie in de evolutie
Door het principe van de assymetrische evolutie, onstaat geleidelijk aan een diversiteit aan "stabiele" evolutiepatronen.
Bepaalde evolutiepatronen krijgen stilaan een zekere "betekenis" voor elkaar of voor de omgeving ; met andere woorden, er onstaat geleidelijk aan informatie in de evolutie. Deze informatie zal meer en meer de selectie in de evolutie gaan beïnvloeden. De aanwezigheid van informatie, onder welke vorm dan ook, betekent dat de evolutie een zekere "voor"kennis heeft en het is onwaarschijnlijk dat de evolutie die daarop volgt, niet door deze voorkennis zal worden beïnvloed.
Sommige van deze evolutiepatronen of interactoren zullen informatie bevatten, die (toevallig) het onstaan van "zelfgelijkaardige" patronen versnelt. Evolutiepatronen die het onstaan van zelfgelijkaardige patronen bevorderen, zullen snel toenemen in aantal.
Deze evolutiepatronen hebben immers een competitief voordeel tegenover de evolutiepatronen die even stabiel zijn, doch deze zelfbevorderende eigenschap niet bevatten. Daardoor zullen ze toenemen in aantal, of met andere woorden, hun "populatie" zal toenemen.
Deze toename of groei van de populatie kan echter niet onbeperkt doorgaan, omdat de M/E/I-onderdelen die deze groei moeten "voeden", beperkt zijn in aantal.
Hierdoor onstaat onvermijdelijk een "competitie" tussen deze zelfbevorderende evolutiepatronen, waarbij enkel de best aangepaste populatie kan blijven groeien. Dit is het principe van de "survival of the fittest" of "het recht van de sterkste".
Het principe van "de blinde evolutie" vormt samen met het principe van de "survival of the fittest", het principe van "de natuurlijke selectie".
We zullen eerst de natuurlijke selectie in de biologie verduidelijken, doch daarna zal aangetoond worden dat natuurlijke selectie ook buiten de biologie, bijvoorbeeld in de pre-biotische evolutie een belangrijke rol heeft gespeeld.
Terug naar de index van dit hoofdstuk
3.1. Natuurlijke selectie in de biologie
In het proces van zelforganisatie en natuurlijke selectie gaan zelforganisatie en natuurlijke selectie hand in hand.
Omdat we in de vorige hoofdstukken zijn reeds dieper ingegaan zijn op de zelforganiserende principes, zullen we in dit hoofdstuk de principes van de natuurlijke selectie diepgaander beschrijven.
Terug naar de index van dit hoofdstuk
Pas bijna decaden na de ontdekking van het principe van "natuurlijke selectie" in de biologie, vatte Darwin zijn ideeën samen in zijn boek "On the origin of species by means of natural selection, or the preservation of races in the struggle for life".
Hij vatte zelf zijn ideeën als volgt samen :
" Omdat er meer individuen ter wereld worden gebracht dan er kunnen overleven, moet er in elk geval een strijd voor het bestaan plaatshebben ; hetzij tussen individuen van dezelfde soort, hetzij met de fysische levensomstandigheden. Is het daarom niet waarschijnlijk dat variaties, die op één of andere manier nuttig zijn voor een individu, zich in de loop van duizenden generaties voordoen ? En als deze zich voordoen, kunnen we er dan aan twijfelen dat individuen met een voordeel - hoe klein ook in vergelijking met andere individuen - de grootste kans hebben op te overleven en zich voort te planten ? Aan de andere kant kunnen we er zeker van zijn dat elke variatie die nadelig is, hoe weinig ook, vernietigd zou worden. Dit behoud van gunstige variaties en het verwerpen van ongunstige variaties, noem ik natuurlijke selectie."
Charles Darwin was slechts 22 jaar toen hij in 1831 inscheepte aan boord van de HMS Beagle, een zeilschip dat een reis rond de aardbol zou maken om de kusten van Zuid-Amerika in kaart te brengen. Vijf jaar later had Darwin’s evolutietheorie reeds vorm gekregen in zijn geest en in zijn notitieboeken. De reden waarom hij zolang wachtte met het publiceren van zijn theorieën was dat hij voldoende bewijsmateriaal wou verzamelen ter ondersteuning van zijn evolutietheorie. Tot in het begin van de negentiende eeuw geloofden de natuuronderzoekers immers in de onveranderlijkheid van de soorten. De Zweedse onderzoeker Linnaeus schreef nog in de eerste helft van de achtiende eeuw : "Er zijn zoveel soorten als in het begin door het oneindige Wezen zijn geschapen".
Binnen een dergelijke wetenschappelijke context was het lanceren van nieuwe evolutieprincipes een gevaarlijke activiteit, die best ondersteund werd door talrijke bewijzen.
Nochtans zijn de ideeën van Darwin niet louter aan zichzelf toe te schrijven, doch zijn ze ook een gevolg van de evolutie van de wetenschappelijke ideeën in die periode.
Nog voor zijn zoektocht naar bewijzen, was Darwin Charles Lyell’s "Principles of Geology" beginnen lezen. Lyell had waargenomen dat het fysische aardoppervlak lange, trage en continue wijzigingen onderging. Op basis van fossiele waarnemingen speculeerde Lyell dat planten en dieren geleidelijk ontstonden, variaties ondergingen en geleidelijk uitstierven. Darwin had ook een essay gelezen dat 50 jaar eerder was gepubliceerd door de econoom Thomas Malthus. In zijn "Essay on the principles of population" bewees Malthus dat populaties een groeipotentieel hebben dat veel hoger is dan kan bijgehouden worden door de benodigde voedingsmiddelen. Malthus besloot hieruit dat organismen, door de relatieve schaarste aan voedingsmiddelen, gedwongen worden een strijd te leveren om te overleven ("Struggle for life") : hierdoor kunnen enkel de sterkste individuen overleven ("Survival of the fittest").
Darwin’s eigen observaties vormden echter de basis voor zijn theorie. In Patagonië vond Darwin gefossileerde beenderen van luiaarden ter grootte van een koe. Ze geleken echter goed op hun nakomelingen die nog steeds Centraal- en Zuid-Amerika bewoonden. Bovendien had Darwin gedurende zijn reizen verregaande variaties gezien in vormen en geografische distributies van planten en dieren. Nergens echter waren de planten en dieren opmerkelijker dan op de Galapagos archipel. Hier ontdekte Darwin reuzeschildpadden en voorhistorisch uitziende leguanen. Zijn bijzonderste waarneming was wellicht dat gelijkaardige diertypes duidelijke variaties vertoonden in lichaamsbouw en functies naargelang het eiland van de archipel. Hij realiseerde zich hierdoor dat er 2 belangrijke factoren zijn in de biologische evolutie : ten eerste is er de "adaptatie" en ten tweede de "isolatie".
De verschillen in populaties hadden te maken met aanpassingen (adaptaties) aan de verschillende omgevingen. "Adaptatie" is in de biologie een genetisch gebonden kenmerk dat resulteert in een individu dat beter is aangepast aan zijn omgeving. Dit kan gaan van bijvoorbeeld kaken die aangepast zijn aan de diverse types voeding, inkapseling om vriestemperaturen te overleven, instinctieve gedragingen om predatoren zoals slangen te vermijden, enz.... Een schoolvoorbeeld zijn de verschillende specialisaties in lichaamsvorm en bektype van de vinken op de Galapagos eilanden (de "Darwin-vinken"). De aanpassingen van de vinken stemmen overeen met het verschillend voedingsgedrag van de vogels. Vinken met een sterke bek eten zaden, deze met een smalle puntige bek eten insecten,... enz.
Darwin redeneerde dat individuen met gunstige aanpassingen meer overlevingskansen hadden dan de onaangepaste individuen, waardoor de aangepaste individuen na enkele generaties het overwicht krijgen in de populatie.
Bovendien zag Darwin in dat organismen geografisch geïsoleerd zijn op de verschillende eilanden. Dit is een selectievoorwaarde voor voortplantingsisolatie. De voortplantingsisolatie is de scheiding van een populatie in groepen die onderling niet paren (en eventueel uitgroeien tot verschillende soorten). In dergelijke geïsoleerde omstandigheden kunnen verschillen in vorm, functie of gedrag zich uitbreiden onder de verschillende individuen van de geïsoleerde groep. Door deze isolatie kunnen zich geleidelijk aan verschillende soorten (species) ontwikkelen uit de geïsoleerde groepen.
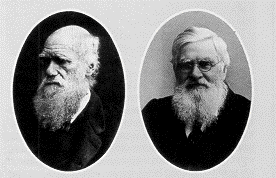
Figuur H8F2 : Charles Darwin (links) en Alfred Russel Wallace (rechts), ontdekkers van de evolutie door "natuurlijke selectie"
Darwin was echter niet de enige wetenschapper die de theorie van de natuurlijke selectie formuleerde. Alfred Russel Wallace, een andere Brit, had eerst planten en dieren bestudeerd in Brazilië en later in Maleisië. Darwin’s verwondering moet groot geweest zijn toen hij een artikel van Wallace ontving, waarin de theorie beschreven stond waarvoor Darwin reeds verschillende jaren bewijzen aan het verzamelen was. Door hun vriendschappelijke banden werd de essentie van hun theorie gezamelijk voorgesteld in London in 1858. Daar waar Wallace de nadruk legde op het competitieaspect, legde Darwin de klemtoon op het reproductieve succes, een concept dat stilaan het basisidee van de evolutieleer werd.
Met zijn "Origin of species" toonde hij aan met gegevens uit de paleanteologie, de vergelijkende anatomie, de embryologie en de dierenaardrijkskunde dat de evolutie als proces een algemeen verschijnsel is bij levende organismen. Gelijktijdig gaf hij voor het evolutieproces ook een heldere verklaring, namelijk de theorie van de natuurlijke selectie.
Terug naar de index van dit hoofdstuk
Toen Charles Darwin zijn bekende boek schreef, erkende hij dat er een serieus bezwaar was tegen zijn theorie. Hoe kan overerving van veranderingen plaatsgrijpen, als iedere spontaan ontstane gunstige verandering in een plant of dier gedoemd was om verdund te worden in de volgende generaties ? Het probleem was dat hij overerving zag als het mengen van kenmerken van de vader en de moeder, wat misschien wortelde in het oude idee dat bloed het erfelijk materiaal bevatte. "Niemand weet waarom een kind in bepaalde kenmerken vaak op de grootvader of grootmoeder lijkt", schreef hij.
Dit probleem werd opgelost door de Oostenrijkse monnik Gregor Mendel (1822-1884).
De verklaring voor Darwins probleem van grootouder en kleinkind ligt in het feit dat de erfelijke informatie (de genen), vrijwel onveranderd van generatie op generatie worden doorgegeven.
Er is geen sprake van mengen of verdunnen, zoals de aanhangers van zachte erfelijkheid dachten. Wel kan het effect van het gen gemaskeerd worden. Zulke genen noemt men recessief. Van andere genen kan het effect niet worden gemaskeerd : deze genen noemt men dominant.
De essentie van de idee van Mendel is dat erfelijkheid niets anders is dan de overdracht van informatie-eenheden (of genen).
Deze genetische informatie in levende systemen, heb ik de "Gamma-replicator" genoemd in het kader van dit boek. Alle biologische wezens (planten, dieren) hebben een "Gamma-replicator", doch ook de andere levende systemen, zoals organisaties en maatschappijen. Ook bij deze laatste is het voortbestaan gewaarborgd door het doorgeven van de informatie die aan de basis ligt van de ontwikkeling van de levende systemen. Ook bij deze systemen zal natuurlijke selectie de ontwikkeling van dominante informatie bevorderen. Het geheel van gemeenschappelijke informatie in de replicatoren van een soort levende systemen, heb ik eerder de "Gamma-attractor" genoemd.
De principes van de natuurlijke selectie, zoals die gelden voor de biologische wezens, gelden ook voor de andere levende systemen. Voor verder ge-evolueerde systemen, zoals menselijke organisaties, zijn echter ook de hogere principes van evolutionaire selectie van toepassing. Hierop wordt verder in dit hoofdstuk teruggekomen.
We zullen echter eerst nog een aantal voorbeelden geven van de natuurlijke selectie bij biologische wezens.
De selectie heeft alleen een onmiddellijk effect op het individu als geheel en niet op het geheel van de genetische informatie (de replicator). Als er eigenschappen zijn die door verschillende genen samen worden bepaald, zullen ook alle betrokken genen samen door de selectie worden beïnvloed. Bij een evolutie van een orgaan (bijvoorbeeld het oog van een zoogdier), zijn vele genen betrokken die de structuur en de functie van alle delen van het orgaan bepalen (netvlies, ooglens en glasachtig lichaam,...) enkel gezamelijk belangrijk voor de selectie. In een vroeg stadium in de evolutie van het oog, biedt een mutatie die leidt tot een verbeterde ooglens op zichzelf geen enkel selectief voordeel. Alleen als het netvlies ook een betere beeldkwaliteit oplevert, zal een dergelijke mutatie de levenskansen doen toenemen.
In bepaalde types selectie is de selectie sterk gedreven vanuit de informatiekenmerken van het levend wezen (bijvoorbeeld bij sexuele selectie), en in andere meer door de kenmerken van de omgeving (bijvoorbeeld selectie in catastrofes) ; een tussenweg vinden we bij selectie door overleving, waar zowel de kenmerken van de omgeving als die van het individu een belangrijke rol spelen. We zullen van deze drie types enkele voorbeelden geven.
In elk geval speelt de semiotische omgeving een belangrijke rol in de "interpretatie" van de informatie in de replicatoren en de Gamma-attractoren van de levende systemen.Zowel Darwin als Wallace begrepen het belang van de omgeving voor de natuurlijke selectie. Geen van beiden echter zag in dat de omgeving als een reservoir van selecterende informatie kon worden aanzien. Het was de in Estland geboren en naar Duitsland ge-emigreerde bioloog Jacob von Uexküll (1864-1944) die de omgeving ("die umwelt") zag als een grootwarenhuis van signalen. Reeds in 1940 schrijft hij romantisch :"Staande voor een weide met bloemen, overvlogen door zoemende bijen, fladderende vlinders, dartele waterjuffers, sprinkhanen die over de grasprieten springen, hollende muizen en langzaam kruipende slangen, stellen we onszelf intuïtief de volgende vraag. Heeft de weide voor elk van deze wezens dezelfde betekenis als diegene die wij eraan geven ?." En zoals vele doorbraken in de wetenschap starten met het stellen van zeer goede vragen, leidt ook deze zeer goede vraag tot een kentering in de evolutiebiologie. Het antwoord op de vraag is immers neen, wat impliceert dat elk dier er een verschillende betekenis aan geeft. Als illustratie van het antwoord op deze vraag in zijn "ümwelt-theorie" geeft hij het voorbeeld van een bloem.
- Een jong meisje ziet een bloem als een decoratief element.
- Voor een mier is de stengel van de bloem een ladder, om tot bij de bloemblaadjes te komen.
- De larven van bepaalde insecten boren gaten in de stengel om materiaal te bekomen voor het bouwen van een "huis" : de bloem is hier dus een voorraad van bouwmaterialen.
- Voor een koe is de bloem een vorm van voeding, net als het gras.
Elk van deze levende wezens geeft dus aan de bloem een betekenis, die past binnen zijn eigen "ümwelt" of context.
Uexküll is hiermee de vader van de biosemiotische visie op de evolutietheorie, tegen zijn eigen wil in. Hij geloofde immers niet in de evolutietheorie zoals die was vooropgesteld door Darwin ! Precies door zijn sceptiscisme tegenover de theorie van Darwin, hebben zijn eigen ideeën aanvankelijk moeilijk ingang kunnen vinden.
In de huidige biosemiotische visie op de evolutie, kunnen de theorieën van Darwin en Uexküll echter perfect verzoend worden, omdat ze complementair zijn.
We zullen daarom nu verder bekijken hoe de informatie in de omgeving, een selecterende invloed uitoefent op de informatiepool in de Gamma-replicatoren en -attractoren van levende systemen.Terug naar de index van dit hoofdstuk
Een groep van dragers van bepaalde informatiekenmerken kan in een beperkt gebied door een besmettelijke ziekte, een onweer, een bosbrand of door andere omstandigheden plots uitsterven. In dat gebied breidt zich dan een populatie uit met een andere genetische samenstelling.Eventueel kunnen dit toevallig overlevende vormen met nadelige mutaties zijn. De toevallige dood of het toevallig overleven kunnen dus van groot belang zijn voor de samenstelling van de populatie. Hoe kleiner de populatie is, hoe waarschijnlijker dat er toevallige verschuivingen in de genfrequenties kunnen voorkomen, doordat bepaalde genen verloren gaan of toenemen in aantal. Deze toevallige schommelingen worden "genetische drift" genoemd. Statistisch kan aangetoond worden dat bij een kleine populatie, een mutatie erg snel kan doorbreken of volledig verdwijnen, ongeacht het feit of de mutatie al dan niet voordelig is.
We kunnen hiervan twee typische voorbeelden geven. Als eerste voorbeeld bekijken we de oorzaken en invloed van massale extincties van diersoorten in de evolutie, en als tweede voorbeeld de invloed van parasieten op populaties van gastheren.
Terug naar de index van dit hoofdstuk
- Massale extincties
Met massale extinctie wordt het uitsterven van een groot aantal levende systemen (planten, dieren,…) bedoeld, gedurende een relatief korte periode (een paar miljoen jaren). Sinds het verschijnen van het leven op aarde zijn er ongeveer een tiental dergelijke periodes van massale extinctie waar te nemen. Bij deze massale extincties zijn een groot aantal soorten dieren betrokken, zowel ter land als ter zee. Om die reden moet de oorzaak ervan een catastrofe zijn die een groot gebied van de aarde treft, en is het onderzoek ervan voornamelijk het terrein van de geologen.
Door het onderzoek van sedimentlagen in de aardbodem, kan men een reconstructie maken van het aardoppervlak op diverse plaatsen van onze aardbol. Uit deze sedimentlagen, en de fossiele resten van planten en dieren, kan men het klimaat in die tijd reconstrueren. Hoewel meerdere factoren aan de basis liggen van deze rampen, heeft men kunnen vaststellen dat deze vrijwel altijd gepaard gaan met zware klimaatwijzigingen.
Eén van de belangrijkste van dergelijke extincties is deze op het einde van de Perm periode, ongeveer 250 à 255 miljoen jaar geleden. Men schat dat toen ongeveer de helft van alle mariene families van de toen bestaande diersoorten is uitgestorven.
Deze periode viel samen met een algemene afkoeling van het klimaat, die als gevolg had dat voornamelijk het leven in de tropen het zwaar te verduren kreeg. De fundamentele oorzaak voor deze extinctie zou te vinden kunnen zijn in de verschuiving van de continenten op aarde. Rond het einde van de Perm-periode lag één van de continenten op aarde, dicht bij de polen. Hierdoor werd op aarde relatief meer zonne-energie weerkaatst, met afkoeling als gevolg.
Ook op het einde van de Krijt-periode (65 miljoen jaar geleden) deed zich een massale extinctie voor, die minder drastisch was dan deze op het einde van het Perm, doch wel meer tot onze verbeelding spreekt. Bij deze extinctie waren voornamelijk de dinosauriërs getroffen, waardoor in de daaropvolgende evoluties voornamelijk de zoogdieren konden doorbreken. De fascinerende dinosauriërs hadden toen nochtans een dominerende positie in het dierenrijk, die ze te danken hadden aan hun goede aanpassing op het heersende ecosysteem, ondanks alle competitie met de toen levende zoogdieren en reptielen. Deze dinosauriërs zijn erin geslaagd gedurende ongeveer 100 miljoen jaar het dierenrijk te domineren. Hun massale extinctie is gevolg van het feit dat ze zich niet snel genoeg konden aanpassen aan de snel wijzigende omstandigheden in hun leefwereld, ten gevolge van de catastrofe.
Een mogelijke verklaring voor deze catastrofe, is de botsing van een enorme komeet of meteoor, met de aarde.
Het uitsterven van planten in deze periodes laat een ander beeld zien. De planten op aarde zijn relatief immuun gebleven voor massale uitsterving. Deze eigenschap danken ze mogelijk aan het feit dat veel planten zich vegetatief of ongeslachtelijk kunnen voortplanten. Hierdoor kan een nieuwe plant of boom onstaan, uit een klein stukje plant. Uit omgevallen bomen kunnen de stronken bijvoorbeeld aanleiding geven tot de ontwikkeling van nieuwe volwassen bomen. Als bij dieren een bepaalde groep uitsterft, neemt een andere dierengroep de niche over, terwijl bij planten bepaalde groepen langzaam uitbreiden ten koste van anderen. De kern van de relatieve ongevoeligheid van planten ligt dus in de manier waarop de Gamma-informatie van generatie op generatie wordt overgedragen.
- Parasieten als motor van evolutie
Parasieten kunnen binnen een populatie van levende wezens ook catastrofes veroorzaken. Denken we maar aan het AIDS-virus, aan infecties die in de middeleeuwen aan massa’s mensen het leven tot een pest hebben gemaakt, enzovoort…
Deze parasieten schakelen hun rivalen niet zozeer uit door een langzame evolutie, met een steeds verdere perfectionering, maar eerder door het ontwikkelen van onvoorspelbare verrassingstactieken, waartegen de gastheer niet is tegen opgewassen en niet of slecht is voorbereid.
De parasieten kunnen hun verrassende strategieën ontwikkelen, omdat hun levensduur veel korter is dan deze van de gastheer. Daardoor heeft de parasiet een heleboel generaties tijd om nieuwe manieren te ontwikkelen om het verdedigingsmechanisme van de gastheer te omzeilen. Als een gastheer nakomelingen voortbrengt die identiek zijn aan zichzelf, vormen deze een gemakkelijke prooi voor de goed aangepaste parasiet, die perfect is afgestemd op de verdediging van de gastheer. Indien de populatie van de gastheer uit genetisch identieke klonen zou bestaan, zou de parasiet deze populatie in één keer kunnen uitroeien.
Zo zijn er inderdaad voorbeelden waarbij een groot deel van de gastheerpopulatie wordt vernietigd.
- De bossen in de Oostelijke Verenigde Staten bestonden voor 1900 voor ongeveer 25% uit Amerikaanse kastanjes, maar tegenwoordig is deze soort als gevolg van een schimmelziekte vrijwel volledig uitgeroeid.
- De builenpest (Zwarte Dood) heeft in de periode van 1334 tot 1351 naar schatting 25 tot 75 % van de bevolking van Europa en Azië uitgeroeid.
- Het AIDS-virus doodt een groot deel van het aantal seropositieven.
Volgens de "hypothese van de rode koningin" is het enige antwoord hierop dat de gastheer zoveel mogelijk nakomelingen produceert, die zoveel mogelijk van elkaar verschillen ; hierdoor moet de parasiet zijn strategie telkens sterk wijzigen. Een goed hulpmiddel hiervoor is de sexuele voortplanting : hierdoor wordt immers gemiddeld 50% van de genetische informatie gewisseld. Dit heeft het nadeel dat ook gunstige variaties kunnen verdwijnen, doch dit nadeel is te verkiezen boven de mogelijke ondergang van de totale populatie. De sexualiteit beïnvloedt en beperkt het selecterend vermogen van parasieten in de omgeving, en beperkt dus de selectie vanuit de omgeving.
De sexualiteit op zich heeft echter nog een belangrijk selecterend aspect, dat samengevat wordt onder de noemer van "sexuele selectie". De sexuele selectie zorgt ervoor dat de partnerkeuze sterk gericht (geïnformeerd) verloopt, en dus ook de overdracht van ontwikkelingsinformatie over de verschillende generaties heen.
Hoewel we zullen zien dat de "sexuele selectie" nog meer uitgesproken een voorbeeld is van geïnformeerde selectie, kan de selectie ten gevolge van catastrofes ook als vorm van "geïnformeerde selectie" worden aanzien. In het eerste voorbeeld over massale extincties hebben we gezien dat klimaatswijzigingen vaak aan de basis liggen van de massale extincties, en dus aan de basis van deze massale "selectieprocessen". Nu is het klimaat een "zelforganiserend systeem", waarbij de klimaatsveranderingen het gevolg zijn van zelf-organiserende Beta-processen en het ontstaan van Beta-informatie. Dat deze klimaatsveranderingen zich soms in relatief korte tijdsperiodes kunnen voltrekken, is een typische eigenschap van zelforganisatie. Immers, voorbij bepaalde kritische punten, wordt een zelforganiserend systeem instabiel en komt er een overgang naar een nieuwe stabiele organisatievorm (bijvoorbeeld een nieuw klimaat).Terug naar de index van dit hoofdstuk
Bepaalde wijzigingen van de replicatorinformatie zullen de overlevingskansen van het levend wezen bevorderen, terwijl andere deze zullen doen afnemen. Als gevolg hiervan ontwikkelen de soorten zich gedurende de evolutie naar een steeds betere adaptie aan de levensomstandigheden.
Bij de levende wezens op aarde kunnen we talrijke voorbeelden vinden van "aanpassingen" die de overleving van de soorten in kwestie hebben bevorderd.
We zullen hier enkele voorbeelden geven.
- Dieren die leven in extreme omstandigheden
Gelijkaardige dieren die in extreme omstandigheden leven, vertonen een lichaamsbouw die aangepast is aan de specifieke omgeving. Zo zijn warmbloedige dieren die in warmere omstandigheden leven dan hun soortgenoten, relatief groter dan hun collega’s die in koudere omstandigheden leven. Bovendien hebben dieren die in koudere gebieden leven, relatief kortere "extremiteiten" (benen, staart, hals, oren...) dan dieren die in warmere gebieden leven.
Beide regels zijn te verklaren door het feit dat de aanpassingen van de dieren in koudere gebieden (groter volume en relatief kleinere extremiteiten), een relatief kleiner warmteverlies tot gevolg zullen hebben, waardoor de overlevingskansen van de dieren met deze aanpassingen groter worden.
Een voorbeeld vinden we bij de vossen.
Figuur H8F3 : een poolvos (links) en woestijnvos (rechts) zijn typische voorbeelden van dieren die aanpassingen vertonen aan de extreme omstandigheden waarin ze leven
De poolvos (Alopex lagopus) heeft korte oren, staart en benen, terwijl de woestijnvos (Vulpes
chama) een langere staart heeft, met grotere oren en langere benen. Door dit verschil in lichaamsoppervlakte, is het warmteverlies afgestemd op enerzijds de poolomgeving en anderzijds de woestijnomgeving. Kenmerkend bij deze dieren is dat ook de kleur van de vacht is aangepast. De poolvos is helemaal wit, waardoor hij beter gecamoufleerd is in de sneeuw, terwijl de woestijnvos een beige kleur heeft, die hem moeilijker zichtbaar maakt in de woestijnomgeving. Ook echter de prooien van de vossen zijn aangepast aan de omgeving. De poolhaas (Lepus timidus arcticus) is eveneens gekenmerkt door een witte vacht en kortere extremiteiten. De witte vacht maakt hem moeilijker zichtbaar voor de vossen, die hun prooi voornamelijk op basis van het zicht waarnemen.
- Vormen en kleuren
Talrijke dieren bezitten vormen die hen beter moeten beschermen tegen hun predatoren, waardoor hun overlevingskansen groter worden.
Een groot aantal dieren bootst als bescherming levenloze objecten na, zoals bladeren en takjes. Dit verschijnsel wordt aangeduid met de term "mimicry". De rupsen van de zijdevlinder kunnen zich uitzonderlijk goed in hun omgeving laten opgaan door hun lichamen de bladstructuur van hun favoriete voedselplanten te laten volgen.
De Zuidamerikaanse bladvis laat zich in de regenwouden op het water van de Amazone drijven en lijkt daarbij sprekend op een met water doordrenkt stuk vegetatie. Zijn lichaam heeft de vorm van een blad en draagt zelfs een bladsteel. Deze vis is een vraatzuchtige rover, die in staat is om een prooi van ongeveer de helft van zijn eigen lichaamslengte te verslinden. Bovendien kan hij zijn kleur en tekening aan die van de ronddrijvende bladeren aanpassen.
Een ander voorbeeld is de wandelende tak, die door zijn bruine of groene schutkleur, nauwelijks te onderscheiden is van zijn omgeving. In elk van deze gevallen biedt de vorm en/of de kleur een competitief voordeel dat de overlevingskansen sterk verbetert.
Ook echter de omgekeerde strategie, namelijk het gebruik van heel opvallende en felle kleuren, kan de overleving van de sommige planten of dieren bevorderen. Vele vlinders zijn heel kwetsbare dieren. Nochtans zijn vele vlinders in de loop van de evolutie gezegend geworden met prachtige patronen en kleuren. Deze evolutie naar opvallendere kleuren, is echter gepaard gegaan met de ontwikkeling van onsmakelijke of zelfs giftige chemicaliën in hun lichaam. De prachtige kleuren zijn daardoor een waarschuwingsteken voor insectenetende vogels, zoogdieren of reptielen, die intussen de associatie hebben gemaakt en geleerd hebben dat ze deze mooi gekleurde insecten beter kunnen vermijden.
Hetzelfde geldt voor de prachtig gekleurde gifpijlkikkers. Deze zwart- en geel gekleurde kikkers scheiden giftige chemische stoffen door hun huid af. Zuid-Amerikaanse indianen vangen de kikkers om deze stoffen te verzamelen en hun pijlpunten mee in te strijken.
Felle kleuren zijn dus in deze gevallen nog sterkere wapens, dan de defensieve schutkleuren die camouflage moeten bevorderen.
Het is dan ook niet verwonderlijk dat sommige dieren zeer felle kleuren gebruiken, om bij de predatoren de idee te wekken dat ze giftige substanties bevatten of op zijn minst onsmakelijk zijn. Zo heeft bijvoorbeeld de dodelijke koraalslang een evenbeeld dat echter niet giftig is, doch vrijwel over een identiek kleurenpatroon beschikt.
Terug naar de index van dit hoofdstuk
- sexuele selectie
Bij vele soorten dieren verschilt het mannetje zo sterk van het vrouwtje, dat het moeilijk te geloven is dat ze tot dezelfde soort behoren.
Ze kunnen variëren in afmetingen, kleur en lichaamskenmerken, waardoor de natuurlijke selectie wordt beïnvloed.Bij één type van de sexuele selectie (de "intrasexuele sexuele selectie"), moeten de mannetjes een fysisch gevecht leveren met elkaar, waarbij alleen de overwinnaar kan paren met het vrouwtje. Het is dit fenomeen dat heeft geleid tot de ontwikkeling van bijvoorbeeld het grote gewei van het mannetjeshert. Elk jaar moet dit hert een groot deel van zijn beschikbare energie stoppen in de vorming van het gewei, en in de ontwikkeling van de spiermassa om het gewei te dragen. De inspanning wordt echter genetisch terugbetaald, doordat het mannetje met het grootste gewei voor nakomelingen kan zorgen.
Zo krijgen ook mannetjeszalmen die uit de oceaan terugkeren om te paren, heel kenmerkende kromme kaken, die hen moet toelaten beter te kunnen vechten met andere mannetjes. Aangezien deze mannetjes kort na het paren zullen sterven, is het geen nadeel dat de vervormde kaken het verzamelen van voedsel bemoeilijken.
Het was Darwin zelf die reeds op het belang van sexuele selectie heeft gewezen. Hij gaf het voorbeeld aan van castratie bij elanden. Als deze castratie vroeg genoeg gebeurt in het leven van het mannetje, resulteert dit in een volwassen mannetje, dat eerder lijkt op een wijfje. Zo hebben gecastreerde elanden geen gewei. Dit is dus ook een teken dat deze wijzigingen een hormonale oorsprong hebben. Bovendien merkte Darwin op dat mannetjes die behoren tot een polygame soort, vaker extreme vormen van sexueel dimorfisme vertonen. De selectie zorgt er immers voor dat die soorten die het meest moeten vechten om te kunnen paren, hier ook het best voor uitgerust zijn.
Naast deze offensieve vormen van sexuele selectie, vinden we ook defensieve vormen terug. De lengte van de manen bij mannelijke leeuwen bijvoorbeeld, biedt bescherming tegen verwondingen door beten in keel en nek.Darwin beschreef ook uitgebreid, en met een vleug verwondering, de moed van diverse mannetjessoorten bij het leveren van deze pre-paringsgevechten, en tevens de verwondingen die ze daarbij oplopen.
Een tweede type van sexuele selectie is meer subtiel, doch zeker niet minder selectief.
Bij dit tweede type van sexuele selectie is vrouwtje actief betrokken bij de selectie van het mannetje. Uit de genetische variatie aan mannetjes, kiest het vrouwtje het meest aantrekkelijke mannetje.
Dit heeft geleid tot vele merkwaardige baltsgedragingen en indrukwekkende verenpracht bij talrijke vogelsoorten.
Voorbeelden van extreem dimorfisme vinden we bij de "paradijsvogels". De mannetjesvogels bezitten een prachtig verenkleed, dat ze rond de paartijd extra ten toon spreiden door het uitvoeren van een paringsdans. Tijdens dit baltsgedrag kan het vrouwtje het mannetje gadeslaan in al zijn glorie, waarbij de aandacht op het prachtige verenkleed wordt beklemtoond door het uitvoeren van een karakteristieke dans, ondersteund door bijhorende baltsgeluiden.

Figuur H8F4 : sexuele selectie heeft geleid tot het prachtige verenkleed van een mannelijke paradijsvogel (links) en een pauw (rechts)
De uitzonderlijk mooie kleur van talrijke tropische vissen die bij koraalriffen leven, is eveneens voor een groot deel te verklaren door het het fenomeen van de sexuele selectie. In deze koraalriffen krioelt het van vissen met unieke en zeer mooie kleurenpatronen en lichaamsvormen. Daardoor is de nood aan duidelijke herkenning van de eigen soort des te groter, maar is ook de competitie om mooiere patronen ten top gedreven.
Ook bij de mens is de partnerkeuze bepaald door elementen van de sexuele selectie. De instinctieve patronen worden echter overschaduwd door cultureel aangeleerde leefwijzen. De genetische informatie die de selectie beïnvloedt, is aangerijkt met informatie die van generatie tot generatie wordt overgedragen. Voorbeelden van rivaliteit tussen "mannetjes" bij de strijd om dezelfde partner (met soms dodelijke gevolgen) kunnen we vrijwel dagelijks in onze krant terugvinden. Bovendien is het ook duidelijk dat vrouwen, bijvoorbeeld door het gebruik van cosmetica en aangepaste kledij, hun aantrekkingskracht op de mannelijke sexe willen verhogen. De sexuele selectie die de mens doorvoert alvorens een partner te selecteren om voor het nageslacht te zorgen, gaat echter veel verder dan "baltsgedrag" en uiterlijke verleiding. Opnieuw kunnen we hiervan dagelijks voorbeelden terugvinden in onze kranten, in de contactadvertenties. Begrippen als vriendelijkheid, intelligentie, academische of andere titels, leeftijd, en dergelijke worden gebruikt om mogelijke partners te selecteren, al dan niet door bemiddeling van een huwelijksbureau.
Het valt bovendien te verwachten dat door de toegenomen mobiliteit, en de toegenomen communicatiemogelijkheden, de partnerkeuze steeds selectiever zal gebeuren, omdat de partners uit een steeds groter aanbod (en variaties) zullen kiezen. Het positieve gevolg hiervan is dat de kans stijgt op het vinden van een geschikte partner. Een ander gevolg is dat, doordat de partners steeds beter bij elkaar zullen passen, en dus wellicht ook meer gemeenschappelijke kenmerken zullen hebben, dat deze sterke selectie zal leiden tot een verdere diversificatie van de families van de menselijke rassen, waarbij de rijken, intellectuelen, mooien, steeds rijker, intellectueler of mooier zullen worden, maar ook het omgekeerde denkbaar is.Terug naar de index van dit hoofdstuk
Natuurlijke selectie is een evolutieproces dat leidt tot steeds verdere adaptie van soorten aan de omgeving. Adaptatie op zich zal echter niet leiden tot de vorming van nieuwe soorten van levende systemen. Nieuwe soorten kunnen enkel ontstaan wanneer een groep wordt afgescheiden, of geïsoleerd van de totale groep waarvan ze deel uitmaken, zodat hun informatiepool door natuurlijke selectie afzonderlijk kan ontwikkelen van deze van de hoofdgroep waarvan ze deel uitmaakten. Er zijn echter verschillende manieren waarop deze isolatie kan worden veroorzaakt.
Eén van de belangrijkste vormen van isolatie, is wellicht de geografische isolatie. Deze kan bijvoorbeeld optreden als gevolg van klimaatsveranderingen. Door het ontstaan van steppen, moerassen of met ijs bedekte gebieden kan het areaal van een populatie in verschillende delen gesplitst worden. Een belangrijke oorzaak van soortvorming op aarde is de continentendrift. Door het uiteendrijven van de continenten op aarde, werden tal van populaties door onoverbrugbare oceanen in geïsoleerde groepen ingedeeld. Zo is bijvoorbeeld de typische fauna en flora van het Afrikaanse continent (met zijn giraffen, diverse types antilopen,...) ontstaan, en is sterk verschillend van de fauna in Australië, die gekenmerkt is door zijn unieke buideldieren.Een andere belangrijke vorm van isolatie is de ecologische isolatie, waarbij ondergroepen een specifieke niche gaan innemen in de omgeving. In Hawai bestaan er twee soorten fruitvliegen, die zo sterk op elkaar gelijken, dat een waarnemer het verschil nauwelijks kan merken. Beide soorten leven van het sap van één welbepaalde boom. Gedurende de evolutie heeft de ene groep fruitvliegen de andere niet verdrongen, doch integendeel zijn twee groepen zich meer en meer gaan specialiseren. De ene soort leeft nu voornamelijk van de sappen hoog in de bomen, terwijl de andere groep zich gespecialiseerd heeft in de sappen dichter tegen de grond aan.
Verschillen in gedrag kunnen leiden tot gedragsisolatie (ethologische isolatie). De merkwaardige baltspatronen van talrijke vogelsoorten, of de eigenzinnige paringsgewoonten van sommige zoogdiersoorten kunnen kruising tussen gelijkaardige individuen bevorderen of verhinderen. Een mooi voorbeeld van gedragsisolatie vinden we terug bij de vuurvliegen. Een vrouwelijke vuurvlieg zal enkel paren met een mannetje, waarvan de nachtelijke lichtpatronen overeenkomen met deze van de soort waartoe ze behoren. De mannelijke vliegpatronen zijn het gevolg van sexuele selectie, die geleid heeft tot een verschillend reproductief succes. Zo heeft de evolutie geleid tot een enorm grote verzameling van visuele, chemische en acoustische signalen die hebben bijgedragen tot de sexuele selectie en een groter voortplantingssucces van bepaalde individuen. Sexuele selectie is dus een belangrijke oorzaak van gedragsisolatie.
Al deze vormen van isolatie maken de bevruchting van eicellen onmogelijk, omdat de zaadcellen de eicellen niet kunnen bereiken of bevruchten.
Een subtielere vorm van isolatie is deze waarbij de eicel wel kan worden bevrucht, doch niet kan ontwikkelen tot een embryo of normaal volwassen organisme. Zo kunnen schapen en geiten bijvoorbeeld met elkaar paren, doch het bevruchte ei kan niet in leven blijven.
Terug naar de index van dit hoofdstuk
3.2. Natuurlijke selectie in de pre- en post-biotische evolutie
De natuurlijke selectie is voornamelijk beschreven voor biologische organismen. Nochtans is ze ook geldig voor andere levende systemen. Ook in de pre-biotische evolutie zijn er evolutiefasen geweest waar de natuurlijke selectie een belangrijk evolutiekenmerk is geweest.
- Het onstaan van de Gamma-replicatoren op aarde
De biologische wezens zijn pas kunnen ontstaan na een fase waarin zelfreplicerende molecules zijn ontstaan. Deze zelfreplicerende molecules zijn onstaan door het proces van "zelforganisatie en natuurlijke selectie", dus door een combinatie van zelforganiserende principes en natuurlijke selectie.
De zelforganiserende principes van het onstaan van deze zelfreplicerende molecules werden reeds in een vorig hoofdstuk beschreven. Hierbij werd aangetoond dat zelforganisatie een belangrijk deelproces is dat geleid heeft tot de vorming van de hypercycli die aan de basis liggen van de eerste Gamma-replicatoren. Deze zelforganiserende processen zijn dikwijls zelfversnellend. Het gevolg hiervan is dat de eerste replicator die vrij goed fungeert, een zeer snelle groei zal kennen, ten nadele van de mogelijke concurrenten. De eerste in zijn soort wint de race, een principe wat soms "survival of the first" wordt genoemd. Dit principe is verantwoordelijk voor het feit dat er veel minder soorten voorkomen, dan oorspronkelijk zou kunnen afgeleid worden uit de principes van de natuurlijke selectie.
Natuurlijke selectie is echter een complementair deelproces waarmee de "beste" zelfreplicerende molecules werden geselecteerd. De molecules die beter geschikt waren voor zelfreplicatie, konden immers beter de aanwezige basismolecules benutten, en daardoor sneller tot replicatie komen. Hierdoor kon het replicatieproces zelf, door natuurlijke selectie, meer en meer verbeterd worden. De molecules die niet tot zelfreplicatie in staat waren, bleven min of meer ongewijzigd in aantal, terwijl deze die aanleiding gaven tot het ontstaan van zelfgelijkvormige molecules, een exponentiële groei hebben gekend.
Door de natuurlijke selectie ("survival of the fittest") worden de interactoren in de evolutie geleidelijk verbeterd. Door het principe van de "survival of the first" ontstaan plotse doorbraken in de evolutie, gevolgd door periodes van zeer snelle evolutie. Doordat beide principes samen werkzaam zijn, leidt dit tot een evolutieproces dat ook wordt aangeduid als "punctuated equilibrium" : periodes van zeer snelle evolutie, worden afgewisseld met periodes van geleidelijke evolutie.- Genetische algorithmen en versnelde ontwikkeling van artefacten
Het bestuderen van de biologie gebeurt traditioneel "in vivo", of "in vitro", door bijvoorbeeld levende cellen te isoleren en verder op te kweken. Recenter zijn de "in silico" studies, of studies door middel van artificieel leven of simulaties van levende systemen in de computer. Eén van de toepassingen van deze "in silico"-studies is het gebruiken van pre-biotische evolutiemechanismen voor optimalisatie van computerprogramma’s.
Het was John Holland die in het midden van de twintigste eeuw, na tegenvallende ervaringen met studies over artificiële intelligentie, het spontane zelforganiserend gedrag dat in de natuur voorkomt, wou simuleren aan de hand van computersimulaties. Na het lezen van een boek over de genetische theorie van de natuurlijke selectie, en met zijn kennis van parallelle computers, komt hij tot het idee van de "genetische algorithmen" voor het ontwikkelen van computerprogramma’s. Holland creëert een electronische genenpool, die bestaat uit kleine subprogramma’s, die elk te beschouwen zijn als een electronisch gen. Elk van deze programma’s heeft de eigenschap voor replicatie en cross-over (kruising van genetische informatie), te vergelijken met wat er in de pre-biotische evolutie is gebeurd.
In één van de experimenten bevatten de initiële subprogramma’s zeer beperkte sorteereigenschappen, voor het sorteren van getallen. Als men deze genenpool aan zijn lot over laat, blijkt dat na enkele generaties, door de mutaties en cross-over in de programmaatjes, er programma’s zijn ontstaan die tot veel efficiënter sorteergedrag komen dan de originele. Zonder ingrijpen van de mens komt men dus tot een verbetering van computerprogramma’s. De principes van de natuurlijke selectie in het pre-biotische periode kunnen dus niet enkel gesimuleerd worden, ze kunnen ook gebruikt worden om sneller nieuwe software te ontwikkelen.
Een andere practische toepassing van deze genetische algorithmen, is de ontwikkeling van nieuwe farmaceutische producten. Hierbij worden in een zware computer de eigenschappen van enkele elementaire organische molecules geprogrammeerd, samen met enkele basisreacties uit de organische scheikunde. Men laat dan de computer, vertrekkend van de basismolecules, nieuwe chemische molecules synthetiseren. Bij elke nieuwe generatie molecules, wordt de hoogste "medische activiteit" van de molecules berekend. De "beste" molecules worden dan geselecteerd om in de volgende generatie tot nieuwe molecules te leiden. In de praktijk blijkt dat na enkele honderden tot duizenden "generaties", er molecules gesynthetiseerd zijn met een hoge (theoretische) medische activiteit. Deze molecules worden dan in het labo gesynthetiseerd, om in vitro en in vivo uitgetest te worden. Op deze wijze kan het gebruik van genetische algorithmen de ontwikkelingstijd van nieuwe farmaceutische producten inkorten.- De eerste materïele deeltjes
Ook nog veel vroeger in de evolutie, namelijk bij het onstaan van de eerste materie, vertoont het evolutieproces kenmerken van de natuurlijke selectie.
Naarmate het heelal afkoelde tijdens de eerste minuten na de oerknal, onstonden achtereenvolgens toevallige associaties van quarks tot hadronen, protonen en neutronen, die zich verder associeerden tot de eerste atomen wanneer de temperatuur verder afkoelde.
Van de vele toevallige associaties die onstonden, werden er slechts enkele "geselecteerd" voor verder gebruik in de evolutie : enkel de stabiele elementaire deeltjes bleven bestaan, terwijl de onstabiele verdwenen. De stabiliteit werd in die fase niet enkel bepaald door de intrinsieke kenmerken van de elementaire deeltjes, maar voornamelijk door omgevingsfactoren als druk en temperatuur. De enorm drastische wijzigingen van temperatuur en druk tijdens de prillen fasen van de evolutie, betekenden letterlijk een zware selectiedruk voor de eerste stabiele deeltjes. Met deze stabiele deeltjes konden verdere associaties ontstaan, die uiteindelijk de bouwstenen gevormd hebben van het heelal zoals we het nu waarnemen. Het recht van de sterkste vertaalt zich in deze evolutieperiode als "het recht van de stabielste". In essentie komt dit echter op het zelfde neer : de best aangepaste en de meest stabiele interactoren zullen voor verdere evolutie geselecteerd worden door de semiotische omgeving, terwijl de minder goed aangepaste en minder stabiele zullen verdwijnen.
Terug naar de index van dit hoofdstuk
De artificiële selectie verschilt van de natuurlijke selectie doordat de mens, en niet de natuurlijke omgeving, de selectiecriteria oplegt en de selectie ook uitvoert. Ze heeft geresulteerd in het onstaan van tal van gedomesticeerde dieren en planten, die door de mens gebruikt worden voor bijvoorbeeld zijn voeding.
Deze selectie is sterk beïnvloed door de kennis en informatie van de persoon die de "veredeling" van de soort aan het doorvoeren is. Nochtans verschilt deze vorm van selectie niet van de natuurlijke selectie op het vlak van de variaties : in beide gevallen zijn de variaties in de Gamma-replicator van de planten of dieren, het gevolg van toevallige wijzigingen.Darwin gebruikte de natuurlijke selectie ook om bepaalde elementen van zijn evolutietheorie te staven. Enerzijds toont de artificiële selectie aan dat de natuurlijke variatie binnen één soort steeds aanwezig is. De mens is in staat om, door het opleggen van nauwkeurig gekozen selectiecriteria, de ingebouwde variaties nog sneller tot uiting te laten komen, en de evolutie van een soort te versnellen.
Anderzijds gebruikte Darwin het feit dat deze soms sterk verschillende gekweekte rassen, nog steeds in staat zijn om met elkaar te kruisen, als bijkomend argument dat de diversiteit der soorten ontstaan is uit gemeenschappelijke voorouders. Slechts na langdurig isolatie, wordt kruising onmogelijk. Dat bijvoorbeeld alle verchillende duivenrassen nog steeds onderling te kruisen zijn, vormde voor hem het bewijs dat ze allemaal afstamden van de gewone huisduif. Ook de talrijke hondenrassen, gaande van Chihuahua’s tot de grote St. Bernardshonden, stammen allemaal af van een gemeenschappelijke wolfachtige voorouder, hoewel ze meer van elkaar verschillen dan bijvoorbeeld vossen van wolven.
Ze zijn het opmerkelijk resultaat van honderden jaren kunstmatige selectie, en tonen aan dat, hoe groter de hoeveelheid informatie is die bij de selectie wordt gebruikt, hoe sneller de evolutie kan gaan. Het is duidelijk dat bij het proces van selectie door overleving, de selectiecriteria minder duidelijk zijn (de selectie is minder goed geïnformeerd), waardoor de evolutie trager gaat. Bij de sexuele selectie daarentegen zijn de selectiecriteria meer uitgesproken, en gaan individuele wezens ook werkelijk de partner selecteren, waardoor de selectie ook meer resultaat oplevert. Bij de artificiële selectie gaan eveneens individuele wezens de selectie bepalen, namelijk de "kwekers" of "veredelaars", die de selectie doen op basis van vaak jarenlange studie en ervaring, waardoor het ook niet verwonderlijk mag zijn dat het resultaat van deze sterk geïnformeerde selecties het meest uitgesproken is. De hoeveelheid en de complexiteit van informatie aanwezig in de semiotische omgeving is dus van cruciaal belang.
De invloed van de informatie op de selectie, werd in één van de eerste hoofstukken besproken in het gedachtenexperiment met de stier en de rode vlag : hoe groter de rode vlag, hoe duidelijker de selectie door de stier. In de voorgaande paragrafen hebben we gezien hoe in de natuur de selectie uiterst subtiel gebeurt, en dat de variatie in mate van geïnformeerd zijn kan variëren van nul bij de puur blinde evolutie, tot zeer sterk bij de artificiële selectie. De essentie van het model met de stier en de rode vlag blijft echter ongewijzigd : hoe duidelijker de informatie, of hoe groter de rode lap, hoe explicieter de selectie.
Terug naar de index van dit hoofdstuk
4. Geïnformeerde variatie in de evolutie
We hebben eerder gezien dat het proces van spontane zelforganisatie, een proces is waarbij
(1) nieuwe informatie wordt gecreëerd en
(2) tegelijkertijd nieuwe variaties ontstaan.
Om die reden kan het proces van zelforganisatie ook aanzien worden als een proces dat "geïnformeerde variaties" doet ontstaan.Samen met het proces van de natuurlijke selectie, leidt dit tot het proces van de "zelforganisatie en natuurlijke selectie".
Een andere vorm van geïnformeerde variaties is door de mens geïntroduceerd.
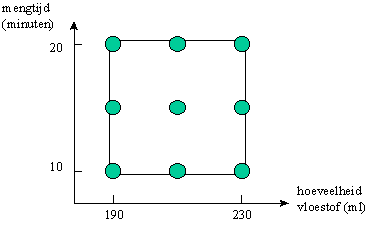
Een situatie die ver afstaat van de toevallige variaties die de natuurlijke selectie kenmerken, vinden we bij de activiteiten van wetenschappers en technologen die onderzoek en ontwikkeling uitvoeren. Beschouwen we bijvoorbeeld een procestechnoloog, die een nieuw proces aan het ontwikkelen is. Hierbij dient bijvoorbeeld een poeder gemengd te worden met een vloeistof. De procestechnoloog wenst op systematische wijze het proces te onderzoeken. Om dit te doen, stelt de onderzoeker een systematische reeks van experimenten op, waarbij hij de te onderzoeken parameters (in dit geval de mengtijd en de hoeveelheid vloeistof), laat variëren tussen 2 relevante waarden. Voor elke parameter worden 3 instelwaarden gekozen, bijvoorbeeld "hoog", "medium" en "laag" (zie grafiek).
Figuur H8F5 : een voorbeeld van een experimentele proefopzet
In dit voorbeeld zal de onderzoeker 9 experimenten uitvoeren (met instellingen overeenkomend met de cirkels in de grafiek), om de optimale combinatie van mengtijd en hoeveelheid vloeistof te kennen. Deze 9 experimenten zijn niet-toevallige, of met andere woorden geïnformeerde variaties, van het proces. Tijdens de experimenten zal de onderzoeker verschillende kenmerken van het proces opvolgen (zoals homogeniteit van het poeder, viscositeit, temperatuur,..). Na verloop van de experimenten zal de onderzoeker de gegevens statistisch verwerken. Hij zal hiermee berekenen met welke procesparameters hij een optimaal resultaat verkrijgt (bijvoorbeeld een welbepaalde viscositeit), waarbij tevens het proces robuust is, en relatief onafhankelijk wordt van toevallige procescondities.
Een dergelijke experimentele proefopzet is een voorbeeld bij uitstek van een geïnformeerde variaties. Aangezien echter de onderzoeker ook een specifiek doel voor ogen heeft, en dus de optimale procesparameters zal kiezen in functie van zijn doelstellingen, is in dit geval ook de selectie geïnformeerd. Deze proef is dus een voorbeeld van wat ik "artificiële constructie" heb genoemd. Aan alle artificiële constructies, gaan echter artificiële variaties vooraf (deze zijn echter niet allemaal even systematisch en doordacht als in dit voorbeeld).
Een zuiver schoolvoorbeeld van artificiële variaties moet inhouden dat de selectie niet geïnformeerd is. In de praktijk komt dit nauwelijks voor. Om die reden zal ik in de volgende paragraaf over artificiële constructie een aantal voorbeelden bespreken, die gekenmerkt zijn door een toenemende hoeveelheid informatie die wordt gebruikt voor de variaties en de selecties.
Terug naar de index van dit hoofdstuk
5. Artificiële constructie in de evolutie
Op aarde is de mens dé expert in het combineren van geïnformeerde variaties met geïnformeerde selectie. Ik heb dit proces "artificiële constructie" genoemd, omdat het meest zichtbare gevolg ervan de constructie van "artefacten" is. Ons dagelijks leven wordt sterk beïnvloed door artefacten als huizen, auto’s, boeken, machines en niet in het minst door abstracte artefacten als de taal en theoretische kennis. In sommige gevallen zijn deze artefacten zeer belangrijk,omdat ze kritische processen in ons leven uitvoeren. Voorbeelden hiervan zijn pacemakers, omdat ze een afwijking proces in het hart kunnen herstellen. Analoog kunnen bepaalde geneesmiddelen zoals insuline, de natuurlijke producten vervangen. Op hogere niveau’s van levende systemen (bijvoorbeeld de organisatie en de maatschappij), verrichten allerlei artefacten levensnoodzakelijke functies. Deze levensnoodzakelijke artefacten, zullen we verder "prothesen" noemen. De mens gaat soms verder in zijn drang naar ontwikkeling en tracht zelfs "artificiële Gamma-replicatoren" te ontwikkelen. Hoewel de mens hier nog niet in geslaagd is, zullen we toch enkele voorbeelden geven van pogingen die in die richting gaan.
Het gebruik van artefacten is echter geen monopolie van de mens. Spinnen bouwen spinnewebben om insecten te vangen ; vogels bouwen nesten om hun jongen tot ontwikkeling te laten komen ; termieten bouwen constructies die als heuvels boven de grond rijzen en bijen bouwen ingenieuze honingraten....
Het onstaan van deze artefacten is echter verlopen volgens een proces van natuurlijke selectie. Spinnen maken immers geen gebruik van experimenten (geïnformeerde variaties) voor het ontwikkelen of optimaliseren van hun spinnewebben en ook vogels experimenteren niet bewust met het bouwen van nesten. De natuur heeft enkel deze spinnen en vogels laten overleven, die webben of nesten konden bouwen die efficiënt genoeg waren voor het vangen van insecten of het opvoeden van de jongen. Het proces van de natuurlijke selectie heeft zeer langzaam geleid tot een steeds beter aangepast zijn van deze "natuurlijke artefacten" aan hun functie, net zoals de vorm en het gedrag van deze dieren zelf is ge-evolueerd door de natuurlijke selectie.
De mens onderscheidt zich door zijn relatief hogere intelligentie, die het proces van geïnformeerde variaties en dus ook geïnformeerde constructie heeft mogelijk gemaakt. Omdat verwerking van Gamma-informatie een cruciale voorwaarde is voor het verwerken van grote hoeveelheden informatie bij de geïnformeerde variaties, kunnen we een sprong zien in de complexiteit van de artefacten, als er een sprong is in de informatieverwerkingscapaciteit. De ontwikkeling van de gesproken taal is wellicht het pronkstuk van de menselijke communicatie, en betekende ook het startschot voor de ontwikkeling van het bewustzijn en de artificiële constructies. De ontwikkeling van de geschreven taal maakte het mogelijk alle ervaringen sneller te communceren en te verspreiden, terwijl de ontwikkeling van een abstracte wiskundige taal, de basis was voor bijvoorbeeld de bloei in de wetenschap in de zeventiende eeuw. Aan de basis van de industriële revolutie in de zeventiende eeuw, ligt precies de ontwikkeling van de wiskundige taal, die gepaard aan wetenschappelijke methodes tot talrijke artefacten heeft geleid.
In de twintigste eeuw heeft de "binaire taal", of de taal van de "enen en nullen" door zijn eenvoud geleid tot het onstaan van de informatica. De software en hardware die uit deze taalrevolutie voortvloeit, doet de aard en het aantal artificiële constructies nog sneller toenemen dan ooit tevoren mogelijk was. De invloed van de informatie op de variaties en selecties in ons dagelijks leven wordt zo groot, dat we dit ofwel over het hoofd zien, ofwel er nauwelijks bij stilstaan.
Omwille van het belang van de "artificiële constructie" voor de evolutie, zullen we achtereenvolgens enkele voorbeelden geven van de constructie van artefacten, de constructie van prothesen en de pogingen om Gamma-replicatoren te contrueren.
- De constructie van artefacten
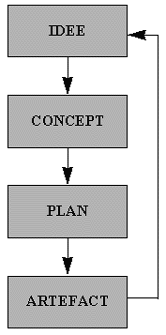
De ontwikkeling van een artefact start meestal met een idee, dat langzaam is gerijpt in de geesten van één of enkele mensen. Een idee dat sinds lang vele geniale en normale geesten heeft geboeid, is de verovering van de ruimte. De beweging van de sterren en vooral de maan hebben de mensheid altijd gefascineerd, wellicht mede omdat de cycli die de zon en maan in relatie tot de aarde maken, ook een belangrijke invloed uitoefenen op het leven op aarde. Het verlangen om de aarde te verlaten, en dichter "bij de goden" te komen, is reeds heel oud. De Egyptenaren slaagden er reeds in de afstand tot de maan in te schatten. Ook hebben vele denkers en fantasten hebben in de loop van de geschiedenis concepten ontwikkeld, waarmee de mens naar de maan zou kunnen reizen.
Figuur H8F6 : artificiële constructie van idee tot artefact
De Griekse schrijver Lucianus beschrijft een "concept" voor een maanreis in één van zijn boeken. In de zestiende eeuw beschrijft de Engelse schrijver en bisschop Godwin, in zijn boek "De man in de maan" meer concreet zijn held met een triomfwagen, in de lucht getrokken door ganzen, ten hemel stijgt en uiteindelijk op de maan landt. Dit werk zal Cyrano De Bergerac inspireren tot talrijke maanfantasieën, waaronder zelfs één waarin naar de zon wordt gevlogen met een meertrapsraket. In de eeuwen die daarop volgen, worden talrijke concepten ontwikkeld voor het reizen naar de maan. Alle concepten zijn beïnvloed door de tijdsgeest, en de stand van de technologie op rond die periode. Edgar Allan Poe beschrijft bijvoorbeeld in 1832 hoe met een luchtballon een reis naar de maan zou kunnen worden gemaakt. Ook Jules Verne beschrijft ballonreizen naar de maan, wellicht geïnspireerd door vertalingen van Poe’s werk. Jules Verne beschrijft ook een reuzekanon van 270 meter lengte, waarmee één van zijn helden naar de maan wordt geschoten.
Concrete plannen voor reële raketten ontstonden pas in onze eeuw, als wetenschap en technologie voldoende matuur zijn om de concepten om te vormen tot concrete plannen. Vooral de militaire industrie, gedreven en gestimuleerd door 2 wereldoorlogen, leidde tot meer concrete realisaties van voornamelijk militaire raketten. Het zou dan maar enkele decennia meer duren, vooraleer de eerste man op de maan "zijn kleine stap voor de mens, doch grote stap voor de mensheid" kon zetten.
Dit voorbeeld van de raket als artefact om de mens naar de maan te brengen, toont precies de kracht van het menselijke denken aan. Ideeën kunnen vrijwel eindeloos veel variaties ondergaan, vooraleer de eerste concreet artefacten te moeten opleveren. De maanreizen hebben eerst millenia van "virtuele evolutie" doorgemaakt, vooraleer de eerste raket is ontwikkeld. Deze raketten hebben talrijke cycli van artificiëke variaties en selecties doorgemaakt, om uiteindelijk tot een plan te leiden dat tot de eerste man op de maan zou leiden.
Het is allemaal minder ambitieus en minder verrijkend begonnen toen de Homo sapiens zijn eerste hut heeft gebouwd, zijn eerste speerpunt heeft ontwikkeld, en andere hulpmiddelen die zijn overlevingskansen hebben vergroot. Sindsdien is het aantal artificiële constructies in stijgende lijn gegaan, is de impact van de natuurlijke selectie op de mens relatief kleiner geworden.
- De constructie van prothesen
Door het ontwikkelen van prothesen is de mens er echter in geslaagd heel expliciet zijn overlevingskansen te verhogen. De beste voorbeelden hiervan zijn de geneesmiddelen, die soms kritische functies van zieke lichamen moesten overnemen. De ontwikkeling van geneesmiddelen verloopt via dezelfde idee tot artefact - cyclus, zoals hierboven geschetst bij de artefacten. De basisidee die hier aan de grondslag ligt is net zo ambitieus als bij de verovering van de ruimte, namelijk het benaderen van de onsterfelijkheid, of meer pragmatisch het verlengen van het leven of verbeteren van de gezondheid. Ook hier is het evolutieproces op zich ge-evolueerd van eerder toevallig (zoals bijvoorbeeld de ontdekking van vaccins) tot sterk geïnformeerd. In de farmaceutische industrie werken dikwijls enkele honderden onderzoekers en medewerkers aan het uitvoeren van artificiële variaties en selecties voor het vinden van nieuwe, en optimaliseren van bestaande geneesmiddelen. Voor het vinden van nieuwe medicijnen worden soms nog processen gebruikt die dicht aanleunen bij de artifiële selectie. Zo worden bijvoorbeeld extracten van (veelal exotische) planten gemaakt, die in feit een groot scala aan mogelijke actieve stoffen bevatten. De variatie in de stoffen is dus niet volledig gecontroleerd. Men gaat op deze extracten of "coctails" van stoffen, zeer expliciete selectiecriteria toepassen, om na te gaan of er geen potentiële geneesmiddelen aanwezig zijn. In andere gevallen zijn de variaties uiterst sterk geïnformeerd. Hierbij construeert men met computers driedimensionele modellen van molecules met mogelijk een geneeskundige activiteit, en simuleert men hoe deze molecules in receptoren van het menselijk lichaam zouden kunnen binden. Door talrijke virtuele variaties, tracht men de molecule te vinden met optimale receptorbinding. Pas dan gaat men over tot de chemische synthese van de molecule. De mens heeft dus geleerd dat het naast elkaar gebruiken van zowel sterk geïnformeerde variatietechnieken, als weinig geïnformeerde variatietechnieken, soms tot het grootste succes kan leiden.
Ook op hogere niveau’s van levende systemen vinden we tal van artefacten en prothesen, gaande van computers, computernetwerken en geheugens ter uitbreiding van het vermogen voor informatieverwerking, tot energiecentrales op basis van kernenergie of waterkracht. Al deze artefacten en prothesen worden continu verbeterd, door opeenvolgende cycli van variaties en selecties. Op deze manier slagen de levende systemen erin zich te verwijderen van het dogma van "aanpassingen aan de omgeving" of "survival of the fitttest", en te komen tot het dogma van de artificiële constructie, dat beter door "aanpassing en creatie van de omgeving" wordt beschreven.
De omgeving wordt dus het lijdend voorwerp van de evolutie, waarvan de talrijke milieuproblemen een stille getuige zijn. De natuurlijke selectie zorgde voor een harmonie tussen levende systemen en omgeving, terwijl bij de artificiële constructie de evolutie van de levende systemen zo snel gaat, dat de omgeving niet de tijd krijgt om zich aan te passen (en soms vice versa).
- constructie van Gamma-replicatoren en Gamma-attractoren
Bij de constructie van artefacten en de constructie van prothesen heb ik extreme voorbeelden gegeven die appeleren aan de droom van de mens om de ruimte te veroveren, en de drang om overleving ad infinitum door te trekken.
De artificiële constructie van Gamma-replicatoren en Gamma-attractoren is op zichzelf gelinkt aan een basiswens van de mens, namelijk het scheppen van het leven op zich. Deze "scheppende kracht" werd eeuwenlang toebedeeld aan talrijke goden, waarvan het de mythes en verhalen zelf onderhevig zijn geweest aan variaties en selecties. Nochtans heeft de mens zelf ook pogingen gedaan om levende systemen, of hun gamma-attractoren te scheppen, of op zijn minst te beïnvloeden of te simuleren. Ik zal hiervan enkele voorbeelden geven.
- Menselijke organisaties en maatschappijen
Misschien het meest succesvolle voorbeeld van levende systemen die door de mens zijn geschapen zijn menselijke organisaties en maatschappijen. Aan de basis van het bestaan van maatschappijen ligt een grondwet (al dan niet geschreven) en zijn er mechanismen om deze grondwet toe te passen. Maatschappijen zijn daardoor de langst levende levende systemen die we op aarde kennen. Zo hebben bijvoorbeeld de Oud-Egyptische en Oud-Griekse maatschappijen eeuwen succesvol overleefd, op basis van hun basiswetten. Hoewel ze gevormd werden door mensen met een relatief korte levensduur, hebben deze maatschappijen in vergelijking met de korte menselijke levensduur een zekere onsterfelijkheid.
Hetzelfde geldt voor menselijke organisaties. Er zijn bijvoorbeeld familiebedrijven, die eeuwen blijven bestaan volgens dezelfde basisprincipes en werkingsmechanismen. De mens staat er vaak niet bij stil dat hij in die situatie deel uitmaakt van het langst levende systeem op aarde.- Genetische manipulatie van planten en dieren
Bij genetische manipulatie wordt rechtstreeks ingegrepen in de genen van planten of dieren : op die manier wordt de informatie van de Gamma-replicator gewijzigd, wat resulteert in een nieuw levend organisme (of een kloon van een bestaand organisme). Deze wijziging van genetische informatie kan opnieuw gebeuren op min of meer geïnformeerde wijze. Zo kan men bijvoorbeeld bacterieën bestralen met radio-actieve stralen. Hierbij zullen de genen van de bacterïen mutaties ondergaan. Vele bacteriën zullen hierdoor sterven, doch enkele kunnen gewijzigde eigenschappen hebben verkregen. In een meer geïnformeerde versie van de genetische manipulatie kan men werkelijk een vreemd gen inbrengen in een bacterie, plant of dier. Zo kan bijvoorbeeld in planten genen inbrengen die de plant resistent maken tegen een niet-selectieve onkruidverdelger (die dus normaal alle planten vernietigt). Op een veld met de genetische gemanipuleerde planten zullen alle planten (inclusief het "onkruid") sterven, met uitzondering van deze planten waarbij de genetische manipulatie succesvol is uitgevoerd.- Artificieel leven
Aangezien er men er niet in slaagt om, startende van nul, nieuwe levende systemen te realiseren, tracht men in de twintigste eeuw meer en meer de computer te gebruiken om het leven niet enkel beter te begrijpen, doch ook het leven te simuleren. Dit virtuele leven wordt aangeduid met de term "artificieel leven".
Eén van de eerste successen op dit vlak was de realisatie van het concept "Life", van de mathematicus John Conway (einde van de jaren 1960). Conway programmeerde een aantal heel eenvoudige regels, die de functie vervulden van de genetische informatie in de Gamma-attractor. Conway’s artificiële wereld bestond uit een "schaakbord" op het computerscherm.
Tot verbazing van de onderzoekers ontstonden snel, op basis van deze regels, eenvoudige patronen van cellen, die een vorm van individueel gedrag begonnen te vertonen. Deze periodieke patronen kregen specifieke namen als "het ship", "de glijder","de kano". De patronen onstonden door een proces van zelforganisatie, na enkele of tientallen generaties. Sommige patronen veranderden van vorm gedurende de verschillende stappen, om daarna terug hun originele vorm te bekomen. Sommige configuraties creëerden na talrijke generaties nieuwe creaties, die ze als het ware uitstootten. Soms werd zelfs een vorm van groepsgedrag tussen de "glijders" waargenomen.
Bovenstaande voorbeelden tonen aan dat de mens wel in staat is Gamma-attractoren te bouwen waarvan hijzelf deel uitmaakt (de organisaties en maatschappijen), doch nog niet in staat is om volledige autonome zelfreplicerende systemen van nul op te bouwen. Zoals de verovering van de ruimte een oude droom is die werkelijkheid is geworden, is de artificiële creatie van Gamma-replicatoren misschien een droom die ook ooit realiteit zal worden...
Terug naar de index van dit hoofdstuk
In dit hoofdstuk werden de variatie en selectie als basiskenmerken van de evolutie verdiept. Er werd aangetoond dat de aanwezigheid of afwezigheid van informatie bij de variatie of de selectie tot verschillende types selectie kan leiden.
Indien zowel de variatie als de selectie niet beïnvloed worden door informatie, kan men spreken van een blinde selectie, omdat dit onvermijdelijk zal leiden tot een blinde evolutie.
Indien de variaties toevallig gebeuren, doch de selectie beïnvloed is door informatie, wordt gesproken van natuurlijke selectie. Aangezien Darwin de grondlegger was van deze evolutiewijze, werd dieper ingegaan op de principes die Darwin, samen met Wallace heeft beschreven. Bovendien werd aangetoond dat de "ümwelttheorie" van Jacob von Uexküll perfect complementair is met de oorspronkelijk gedefinieerde theorie van Darwin en Wallace : de omgeving is een vrijwel oneindig reservoir van informatie, dat door haar rijkdom aan informatie, als selectiemedium bij uitstek fungeert in de evolutie.
Bijzondere aandacht werd ook besteed aan de sexuele selectie, aangezien deze de vorm en het gedrag van talrijke planten en dieren op onze aarde sterk heeft beïnvloed. De artificiële selectie, of de veredeling van planten en dieren door de mens, heeft uiteindelijk geleid tot talrijke gedomesticeerde groenten en fruit die we vrijwel dagelijks consumeren, en tot de diverse huisdieren die ons leven kenmerken. Er werd echter ook aangetoond dat de natuurlijke selectie een evolutieproces is dat niet enkel beperkt is tot de evolutie in de biologie, doch ook een belangrijke invloed heeft gehad op de pre-biotische evolutie en zelfs bij het ontstaan van bijvoorbeeld de eerste materiële deeltjes.Natuurlijke selectie is echter een proces dat moeilijk kan losgekoppeld worden van de zelforganisatie die spontaan optreedt in gradiëntrijke omgevingen zoals onze door zonne-energie overladen aardbol. Om die reden wordt gesproken van het evolutieprincipe van de "zelforganisatie en natuurlijke selectie". Door de samenwerking tussen beide ontstaat een synergistisch effect, doordat zowel variaties als selecties geïnformeerd kunnen gebeuren.
Naarmate meer en meer informatie in onze cultuur doordringt, maken ook de blinde variaties meer en meer plaats voor artificiële variaties. Bij het uitvoeren van wetenschappelijke experimenten zal men bijvoorbeeld kunstmatig variaties inbrengen in de proefopzet, om zo duidelijke conclusies uit de experimenten te kunnen "selecteren". Deze artificiële variaties zijn de basis van de artificiële constructies, of de "constructie van artefacten". Levende systemen zoals de mens, de menselijke organisaties en de maatschappijen, zijn ingebed in een breed gamma van al dan niet levensnoodzakelijke artefacten, die de overleving van het levend systeem moeten bevorderen.Uiteindelijk werd hiermee aangetoond dat evolutieprincipes op zichzelf onderhevig zijn aan evolutie, en dat er een trend is in de richting van meer geïnformeerde evolutieprincipes. Daar waar bijvoorbeeld de eerste levende systemen door de natuurlijke selectie meer en meer aangepast werden aan de omgeving, wordt bij de artificiële constructie de omgeving meer en meer aangepast aan het levend systeem.
De algemene evolutieprincipes zijn schematisch nog eens gesitueerd in onderstaande overzichtsfiguur.
Terug naar de index van dit hoofdstuk