
Persistence:
- Persistent Chlamydial Development:
Persistence
describes a long-term association between Chlamydiae and their host cell
in which these organisms remain in a viable but culture-negative state.
Persistence
may represent a deviation from the typical development of Chlamydiae, resulting
in delayed intracellular growth under the influence of exogenous factors
that may not be as "typical" as cell culture growth conditions. These conditions
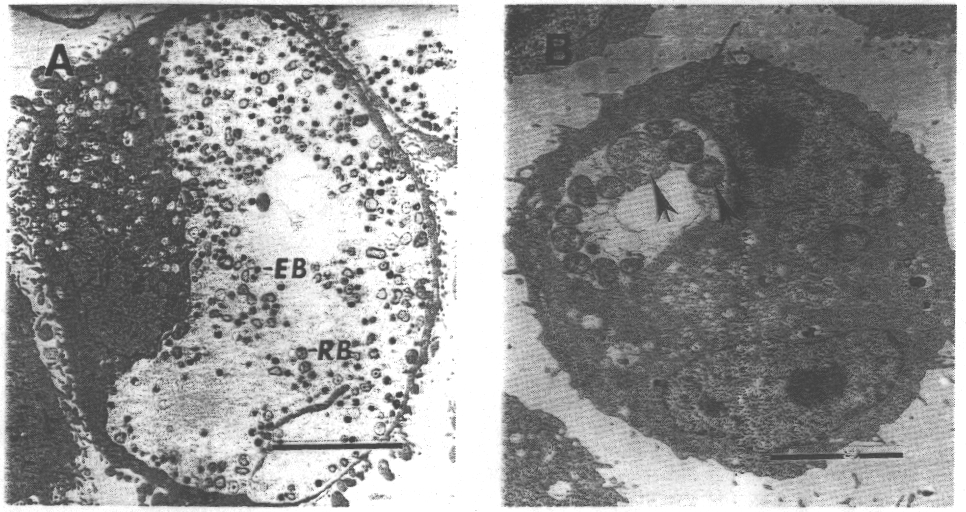
generally delay reticulate body (RB) maturation, inhibit differentiation
to infectious elemental body (EB), and are associated with gross morphological
alterations of RBs typified by markedly enlarged, atypical chlamydial forms.
Delays in chlamydial development in response to exogenous factors suggest
an innate ability of these organisms to persist intracellulary in a unique
developmental form.
Chlamydial persistence has been established in a variety of cell culture
systems; however, conclusive documentation of persistent Chlamydiae in
natural infections remains to be demostrated.
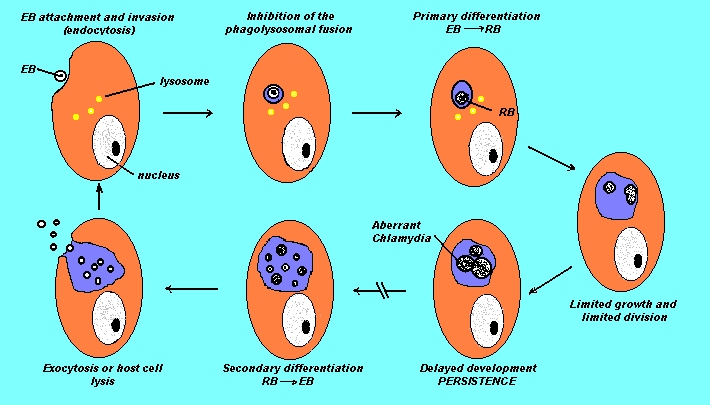
Altered
Chlamydial Development.
*Persistent infections
in cell culture:
Nutrient deficiency-induced persistence:
Chlamydiae are able to invade the deficient host cells and upon entry remain
in a noninfectious but viable state. Addition of complete medium stimulates
growth and recovery of infectious Chlmaydiae.
Depletion of cysteine interrupts chlamydial RB-to-EB differentiation
in ten different serovars of C. trachomatis. This effect is reversible,
with resumed differentiation to infectious forms, upon the addition of
cysteine. Deficiency of other amino acid have a little or no effect
on chlamydial development. These observations suggest that an alteration
in growth and differentiation arises via the requirement for this amino
acid for the biosynthesis of three cysteine-rich proteins (MOMP, 12 and
60 kda.) needed for RB-to-EB differentiation.
Some studies show that intracellular development of C. trachomatis serovars
E and L2 in a medium lacking thirteen amino acids result on reduced infectious
yield, with the development od enlarged abnormal chlamidial forms. This
effect is reversible, with a substantial recovery of infectivity upon the
reintroduction of amino acids.
In conclusion, under conditions in which the host´s soluble pools
for metabolic needs become limited, Chlamydiae may fail to successfully
compete for macromolecular precursors and hence may enter a state of arrested
growth.
Antimicrobial agents and persistence:
Treatment with penicillin has no effect on initial differentiation
of the infecting EB to the RB but prevents the process of binary fission
inducing the development of enlarged, morphologically abnormal chlamydial
forms with a block in the production of progeny EBs. These aberrant forms
become progressively larger with continued culture in the presence of the
antibiotic. Because Chlamydiae are deficient in peptidoglycan the mechanism
of chlamydial growth inhibition by penicillin is unknown. The penicillin
have no effect on the synthesis of the MOMP but the synthesis of the cysteine-rich
60 kda protein is completely inhibited. Removal of penicillin from the
extracellular culture medium results in extensive budding and internal
subdivision of the aberrant chlamydial forms, producing typical RBs with
maturation to infectious EBs.
Ampicilin
also
blocks the conversion of RBs to EBs. The effect of ampicillin on Chlamydia-specific
protein synthesis reveals that arrest of chlamydial development is accompanied
by a marked inhibition in the expression of both 60 kda and 12 kda cysteine-rich
proteins, while the production of MOMP is only minimally affected.
Chloranfenicol
and
chlortetracycline
have
also been shown to interrupt the intracellular development of Chlamydiae.
The effect of these inhibitors of prokaryotic protein synthesis is dependent
on the stage during infection at which the Chlamydia-infected cells are
exposed to these antibiotics. Addition of these inhibitors early in infection
prevents primary differentiation of EB to RB ( persistence is not induced
), whereas exposure later in infection interrupts RB division and secondary
differentiation.
A nearly identical situation occurs when the effect of Erythromycin
on chlamydial development is analyzed. This antibiotic not only inhibits
the RB-to-EB differentiation but also induces smaller inclusions containing
RBs of approximately twice the diameter of typical RBs. Erythromycin binds
to the 50s subunit of the ribosome and may reduce RB ribosomal activity
and subsequent protein synthesis.
Antibiotics that inhibit nucleic acid synthesis also have been shown to
inhibit chlamydial development. 5-fluorouracil affects late stages
of the C. trachomatis LGV growth cycle. Treatment of 48 hours shows smaller
inclusions containing only a few noninfectious larger particles present.
Hydroxyurea,
also blocks secondary differentiation, as before, accompanied by the inhibition
of the synthesis of cysteine-rich proteins.
Sulfonamides as trimethoprim and sulfomethazole at levels
slightly below the MIC inhibit RB-to-EB differentiation characterized by
the presence of gross changes in RB formation.
Immunologically induced persistence:
The most of the
studies about persistence are focus in the T-cell product gamma interferon
(IFN-g)
using HeLa 229 cells infected with C. trachomatis serovar A. Chlamydiae
were among the first nonviral pathogens reported to induced IFNs and were
shown to exhibit growth inhibition in their presence. IFN-g
affects human host cells in vitro by inducing indoleamine 2,3-dioxygenase
(IDO), a nonconstitutive enzyme that catalyzes the initial step in the
degradation of tryptophan to N-formylkynurenine and kynurenine.
Persistence
of C. trachomatis occurs when very low levels of IFN-g
are
added to culture systems following infection of the host cells. This effect
is reversible upon IFN-g
removal from the culture medium. High levels of it inhibit completely chlamydial
development.
IFN-g-induced
persistent organisms not only exhibit a highly unusual intracellular morphology
but also display differential expression of key chlamydial antigens with
continued synthesis of hsp-60, an immunopathological antigen, and reduced
synthesis of MOMP, aprotective antigen. Besides, these forms exhibit a
reduction in the levels of other structural constituents of Chlamydiae,
including the 60 kda envelope protein and lipopolysaccharide.
The effect of
exogenous tryptophan on IFN-g-induced
persistent development revealed that the presence of superphysiological
concentrations of tryptophan interrupted IFN-g-mediated
persistence, resulting in the development of typical chlamydial inclusions.
Cycloheximide, an inhibitor of host protein synthesis, have the same effect
of exogenous tryptophan on IFN-g-mediated
persistence.
Other potential mediators of persistence:
Treatment of
C. trachomatis L2-infected cells with AMPc reversibly interferes
with the correct progression of Chlamydiae through the developmental cycle.
Inclusions are small and immature and remain noninfectious in the presence
of AMPc.
Verapamil,
a Ca2+ antagonist, interferes with chlamydial development inhibiting RB-to-EB
differentiation.
- Persistence of Chlamydiae in vivo:
There are evidence of it in:
Ocular infections:
Although a clear
correlation between trachoma and the infection with C. trachomatis
has been established, Chlamydiae cannot be identified by tissue culture
or immunocytological methods in over 20% of cases, even in the presence
of severe abnormalities.
Older individuals
who have been exposed to repeated infections display complications of ocular
scarring and fibrosis, corneal trauma, and blindness, even though C. trachomatis
is rarely isolated from the disease tissue.
The presence
of a cryptic form of Chlamydiae in the infected tissues is suggest in studies
of individuals who left areas where trachoma was endemic and had
no experienced active disease since childhood but developed acute trachoma
several decades later. Therefore, viable Chlamydiae may be present in a
latent , nonreplicating form, contributing to the progression of disease
toward blinding trachoma.
Genital infections:
The identification
of C. trachomatis as an etiologically agent of infertility in women is
well recognized. Infection of the cervix by C. trachomatis may be chronic,
persisting unrecognized for months to years unless a symptomatic infection
of a male partner occurs.
Approximately
70% of women with C. trachomatis cervicitis have only mild symptoms or
remain asymptomatic. Unrecognized infection can progress, ascending to
the salpinges and fallopian tubes and resulting in pelvic inflammatory
disease.
Chlamydial antigens
have been detected in endometrial and tubal specimens of culture-negative
infertile women.
Chlamydia-specific
DNA has been identified in endocervical cells and fallopian tube tissue
of women with tubal occlusion.
In conclusion,
affected individuals may harbor a persistent form of Chlamidiae that possesses
the characteristics of noncultivatable RBs but retains viability and the
capacity to stimulate immunophatologic changes. However, evidence of chlamydial
antigen and nucleic acids in the absence of cultivable organisms only suggests
the possibility of persisting Chlamydiae.
References:
1. Allan, I., T. P. Hatch, and J. H. Pearce. 1985. Influence of cysteine deprivation on chlamydial differentiation from reproductive to infective life-cycle forms. J. Gen. Microbiol. 131:3171-3177.
2. Allan, I., and J. H. Pierce. 1983. Amino acid requirements of strains of Chlamydia trachomatis and C. psittaci in Mc Coy cells: relationship with clinical syndrome and host origin. J. Gen. Microbiol. 129:2001-2007.
3. Barbour, C. E., K. Amato, T. Hackstadt., L. Perry, and H. D. Caldwell. 1982. Chlamydia trachomatis has penicillin-binding proteins but not detectable muramic acid. J. Bacteriol. 151:420-428.
4. Beatty, W. L., G. L. Byrne, and R. P. Morrison. 1993. Morphological and antigenic characterization of IFN-G mediated persistence Chlamydia trachomatis infection in vitro. Proc. Natl. Acad. Sci. USA. 90:3998-4002.
5. Beatty, W. L., T. A. Belanger, A. A. Desai, R. P. Morrison, and G. I. Byrne. 1994. Tryptophan Depletion as a Mechanism of Gamma IFN-Mediated Chlamydial Persistence. Inf. and Immunity. 62-9:3705-3711.
6. Cevenini, R., M, Donati, and M. La Placa. 1998. Effects of penicillin on the synthesis of membrane proteins of C. trachomatis LGV2 serotype. FEMS Microbiol. Lett. 56:41-46.
7. Hammerschlag, M. R., and J. C. Vuletin. 1985. Ultrastructural analysis of the effect of trimethoprim and sulphamethoxazole on the development of C. trachomatis in cell culture. J. Antimicrob. Chemother. 15:209-217.
8. Oriel, J. D. 1986. The carrier state: C. trachomatis. J. Antimicrob. Chemother. 18SA:67-71.
9. Schachter, J. 1978. Chlamydial infections. N. Engl. J. Med. 298:428-435.
10. Shepard, M. K., and R. B. Jones. 1989. Recovery of Chlamydia trachomatis from endometrial and fallopian tube biopsies in women with infertility of tubal origin. Fertil. Steril. 52:232-238.
11. Taylor
H. R., P. A. Rapoza, S. West, S. Johnshon, B. Munoz, S. Katala, and B.
B. O. Mmbaga. 1989. The epidemiology of infection in trachoma. Invest.
Ophthalmol. Visual Sci. 30:1823-1833.
