
Persistencia:
- Desarrollo Clamidial Persistente:
* Persistencia en
Cultivos Celulares.
- Persistencia inducida por déficit de nutrientes.
- Persistencia y los agentes antimicrobianos.
- Persistencia inducida inmunologicamente.
- Otros mediadores potenciales de persistencia.
* Persistencia en vivo.
Hay evidencia de ella
en:
- Infecciones oculares.
- Infecciones genitales.
- Desarrollo Clamidial Persistente:
Persistencia describe una asociación a largo plazo entre las Clamidias y la célula huésped en la cual estos organismos permanecen viables pero en estado de cultivo negativo. Persistencia puede representar una desviación del ciclo de desarrollo típico de las Clamidias resultando en un crecimiento intracelular demorado bajo la influencia de factores exógenos. Estas condiciones generalmente demoran la maduración del cuerpo reticulado (CR), inhiben la diferenciación a cuerpo elemental (CE) y son asociados con alteraciones morfológicas de los CR´s. La persistencia es caracterizada por el desarrollo de grandes y no infecciosas formas aberrantes que muestran alteración de las principales proteínas clamidiales. Muestran una disminución en los niveles de LPS, OMP 60kda, MOMP y HSP 60kda. Demoras en el desarrollo clamidial en respuesta a factores exógenos sugieren una innata habilidad de este organismo a persistir intracelularmente en una única forma de desarrollo. El estado biológico de las Clamidias y de los factores que llevan a la persistencia en vivo son desconocidos, no obstante, varios estudios han establecido sistemas de cultivo celular para definir parámetros asociados con el crecimiento y la inducción de infecciones clamidiales persistentes.
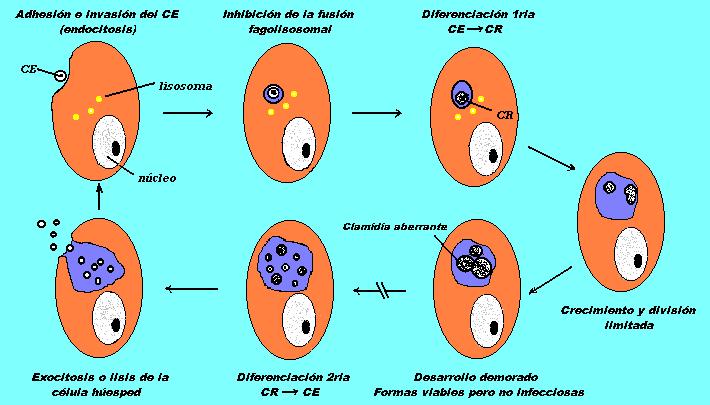
Desarrollo
Clamidial Alterado.
*Persistencia
en cultivos celulares:
Persistencia inducida por déficit de nutrientes:
Las Clamidias son capaces de invadir células en un medio de nutrientes
deficientes y al entrar permanecer en un estado viable pero no infeccioso.
El agregado de medios completos estimulan el crecimiento y recuperación
de Clamidias infecciosas. Se ha observado que la disminución del
nivel de cisteína interrumpe la diferenciación de
CE a CR en diez distintos serotipos de C. trachomatis. Efecto reversible
al agregar cisteína. Deficiencia de otro aminoácido tiene
poco o ningún efecto sobre el desarrollo clamidial. Esto sugiere
que una alteración en el crecimiento o diferenciación es
debida a la falta de cisteína para la biosíntesis de tres
proteínas ricas en cisteína (MOMP, 12 y 60 kda.) necesitadas
para la diferenciación de CR a CE. Se vió en el serotipo
E y L2 de C. trachomatis que en un medio donde escasean los trece aminoácidos
hay bajo rendimiento infeccioso con el desarrollo de grandes y anormales
formas clamidiales. Este efecto es reversible, con una recuperación
substancial de la infectividad al reintroducir los aminoácidos.
En resumen, los factores que comprometen la capacidad biosíntetica
del huésped tienen un efecto mayor sobre la competición por
precursores macromoleculares y entrar así en un estado de persistencia.
Persistencia y los agentes antimicrobianos:
El tratamiento con penicilina no tiene efecto en la diferenciación
inicial del CE al CR pero previene el proceso de fisión binaria
induciendo el desarrollo de alargados, morfológicamente anormales
formas clamidiales con un bloqueo en la producción de CE´s.
Los CR son progresivamente más grandes con cultivos continuados
en la presencia del antibiótico. Como las paredes de las clamidias
son deficientes en peptidoglicano, el mecanismo de la inhibición
del crecimiento por penicilina es desconocido. La remoción de la
penicilina del medio de cultivo lleva a la producción de CR´s
típicos con la posterior maduración a CE´s infecciosos.
Células infectadas con C. trachomatis (LGV) y tratadas con penicilina
mostraron que ésta no tiene efecto sobre la MOMP pero si inhibe
la síntesis de la proteína de 60 kda.
Tratamientos con ampicilina mostraron que ésta bloquea la
diferenciación 2ria de CR a CE induciendo persistencia y que
tuvo un efecto mínimo sobre la MOMP pero una marcada inhibición
de la síntesis de la proteína de 60 y 12 kda.
El cloranfenicol y la clortetraciclina son inhibidores de la síntesis
de proteínas procarióticas, son dependientes de la fase durante
la infección en la cuál las células infectadas con
Clamidias son expuestas a los antibióticos. El agregado de estos
al comienzo de la infección previene la diferenciación 1ria
de CE a CR por lo que no se induce persistencia, mientras que la exposición
tardía en la infección interrumpe la diferenciación
2ria.
La eritromicina bloquea la diferenciación 2ria de CR a CE
induciendo la formación de inclusiones más pequeñas
de lo normal con CR´s de aproximadamente dos veces el diámetro
de CR´s típicos. Este antibiótico se une a la subunidad
50s del ribosoma y reduce la actividad ribosomal del CR y la consecuente
síntesis de proteínas.
También se ha observado que antibióticos que inhiben la síntesis
de ácidos nucleicos inhiben el desarrollo Clamidial induciendo persistencia.
Este es el caso del 5-fluorouracil y de la hidroxiurea.
Se ha observado
que la inducción de persistencia por antibióticos se da con
concentraciones de estos menores a la CIM y que una vez que estos se quitan
del medio de cultivo se produce la reactivación de organismos infecciosos
viables.
En conclusión,
hay un gran número de antibióticos que inhiben la diferenciación
Clamidial y sus efectos parecen ser dependientes del estadío
del ciclo durante el cuál las células infecciosas son expuestas
a estas sustancias. Se ve que el blanco molecular primario para cada antibiótico
es muy amplio lo que lleva a pensar que no es un solo mecanismo por el
cuál se bloquea la diferenciación y se produce la persistencia.
Persistencia inducida inmunologicamente:
La mayor cantidad
de estudios sobre este tema se basan en el estudio del interferón
gama (IFN-g)
que es un producto de los linfocitos T. Las Clamidias fueron los primeros
patógenos no virales que fueron reportados como inductores de interferones
y que inhiben el crecimiento en su presencia. El IFN-g
afecta la célula huésped humana in vitro por la inducción
de indolamina 2,3 dioxigenasa (IDO), enzima que cataliza el paso inicial
en la degradación del triptofano.
La mayoría
de los experimentos sobre el IFN-g
se realizaron usando las células HeLa 229 infectadas con Clamidias
trachomatis serotipo A. Se observa que altos niveles de IFN-g
restringen completamente el crecimiento, mientras que niveles relativamente
bajos del mismo inducen el desarrollo de formas morfológicamente
aberrantes. Se obtienen organismos infecciosos viables luego de la eliminación
del IFN-g.
El efecto de
la cicloheximida, un inhibidor de la síntesis de proteínas
del huésped, sobre células infectadas con formas aberrantes
producidas por IFN-g
muestra que interrumpe el desarrollo del estado de persistencia. Algo similar
ocurre cuando se le agrega triptofano en lugar de cicloheximida.
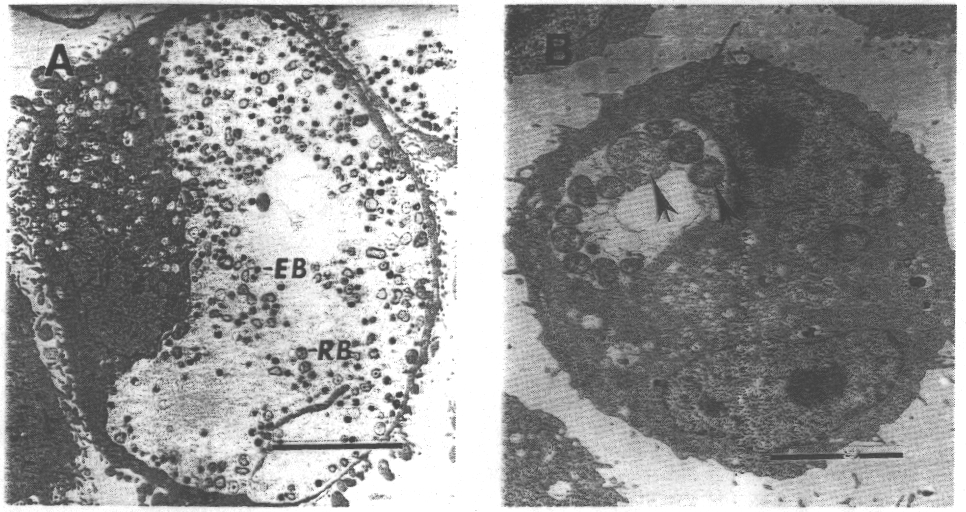
Célula
con una inclusión normal (izq.) y célula tratada con IFN-g
(der.)
Otros mediadores potenciales de persistencia:
Células
infectadas con C. trachomatis L2 tratadas con AMPc mostraron la
interferencia reversible del ciclo de desarrollo normal. Se vieron inclusiones
pequeñas, inmaduras y que permanecieron no infecciosas en presencia
de AMPc.
El verapamil,
un antagonista del Ca2+ que parece ser importante para el crecimiento y
desarrollo clamidial, inhibe la diferenciación 2ria de CR a CE.
* Persistencia en vivo:
Hay evidencia de ella en:
A pesar de la
clara relación establecida entre el tracoma y la infección
clamidial, estas no pueden ser detectadas por cultivo de tejidos o por
otros métodos inmunocitológicos en más de un 20% de
los casos.
Individuos de
edad avanzada quienes han sido expuestos a infecciones repetidas muestran
complicaciones como fibrosis, trauma corneal, ceguera a pesar de que la
Clamidia es raramente aislada del tejido enfermo.
La presencia
de una forma críptica de Clamidia en los tejidos afectados es sugerido
por estudios de individuos que dejaron áreas donde el tracoma era
endémico y no había experimentado enfermedad activa desde
la niñez pero desarrollaron tracoma severo varias décadas
después.
Resumiendo,
las Clamidias podrían persistir en una forma latente y no replicativa,
contibuyendo a la progresión de la enfermedad hacia la ceguera causada
por tracoma.
La identificación
de C. trachomatis como un agente etiológico de la infertilidad en
mujeres esta bien reconocido. Aproximadamente 70% de las mujeres con infecciones
de C. trachomatis tienen moderados síntomas o permanecen asintomáticas.
Infecciones no reconocidas pueden progresar ascendiendo a las trompas de
fallopio y resultando en una enfermedad inflamatoria pélvica. Infecciones
silenciosas son las causas principales de infertilidad con 1/3 de
las mujeres infértiles teniendo una historia de enfermedad inflamatoria
pélvica.
Aunque las Clamidias
son difíciles de cultivar, la presencia de Ag clamidiales y de ácidos
nucleicos son indicadores de organismos persistentes. Estudios han identificado
Ags clamidiales en endometrio de mujeres infértiles y ADN clamidio-específico
en células de cuello y tejido de las trompas de fallopio de mujeres
con oclusión de las mismas.
En conclusión,
individuos afectados podrían dar un hábitat a formas persistentes
que poseen características de CR´s no cultivables pero retienen
la viabilidad. No obstante, la presencia de Ag clamidial y ácidos
nucleicos en la ausencia de organismos cultivables solo sugiere la posibilidad
de Clamidias persistentes.
Referencias:
1. Allan, I., T. P. Hatch, and J. H. Pearce. 1985. Influence of cysteine deprivation on chlamydial differentiation from reproductive to infective life-cycle forms. J. Gen. Microbiol. 131:3171-3177.
2. Allan, I., and J. H. Pierce. 1983. Amino acid requirements of strains of Chlamydia trachomatis and C. psittaci in Mc Coy cells: relationship with clinical syndrome and host origin. J. Gen. Microbiol. 129:2001-2007.
3. Barbour, C. E., K. Amato, T. Hackstadt., L. Perry, and H. D. Caldwell. 1982. Chlamydia trachomatis has penicillin-binding proteins but not detectable muramic acid. J. Bacteriol. 151:420-428.
4. Beatty, W. L., G. L. Byrne, and R. P. Morrison. 1993. Morphological and antigenic characterization of IFN-G mediated persistence Chlamydia trachomatis infection in vitro. Proc. Natl. Acad. Sci. USA. 90:3998-4002.
5. Beatty, W. L., T. A. Belanger, A. A. Desai, R. P. Morrison, and G. I. Byrne. 1994. Tryptophan Depletion as a Mechanism of Gamma IFN-Mediated Chlamydial Persistence. Inf. and Immunity. 62-9:3705-3711.
6. Cevenini, R., M, Donati, and M. La Placa. 1998. Effects of penicillin on the synthesis of membrane proteins of C. trachomatis LGV2 serotype. FEMS Microbiol. Lett. 56:41-46.
7. Hammerschlag, M. R., and J. C. Vuletin. 1985. Ultrastructural analysis of the effect of trimethoprim and sulphamethoxazole on the development of C. trachomatis in cell culture. J. Antimicrob. Chemother. 15:209-217.
8. Oriel, J. D. 1986. The carrier state: C. trachomatis. J. Antimicrob. Chemother. 18SA:67-71.
9. Schachter, J. 1978. Chlamydial infections. N. Engl. J. Med. 298:428-435.
10. Shepard, M. K., and R. B. Jones. 1989. Recovery of Chlamydia trachomatis from endometrial and fallopian tube biopsies in women with infertility of tubal origin. Fertil. Steril. 52:232-238.
11. Taylor
H. R., P. A. Rapoza, S. West, S. Johnshon, B. Munoz, S. Katala, and B.
B. O. Mmbaga. 1989. The epidemiology of infection in trachoma. Invest.
Ophthalmol. Visual Sci. 30:1823-1833.
