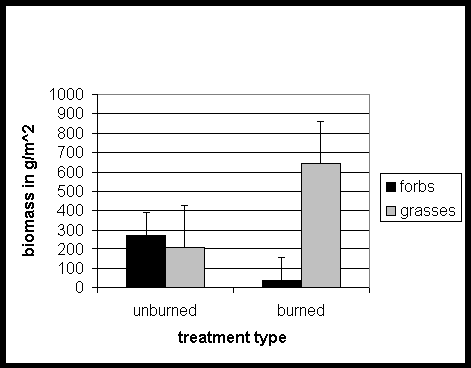
Figure 1: Comparison of the ratio of forb biomass to grass biomass in the unburned and regularly burned plots in tallgrass prairie at Boulder, Colorado.
Kimberly J. Reever Morghan
One Shields Avenue
Department of Agronomy and Range Science
University of California
Davis, California 95616
(530) 752-1701 Phone
(530) 752-4361 FAX
kmorghan@ucdavis.edu
KIMBERLY J. REEVER MORGHAN 1
TIMOTHY R. SEASTEDT
AND
PENELOPE J. SINTON 2
Institute for Arctic and Alpine Research
and
Department of Environmental, Population and Organismic Biology
University of Colorado Boulder, Colorado 80309
Ā
Abstract. Tallgrass prairie ecosystems are maintained as grasslands by regular burning, but burning has been implicated as a potential contributor to invasion by exotic species. We propose that it is not fire itself that creates the potential for invasion, rather it is infrequent fire that creates opportunities for invasion by creating a pulse of nitrogen availability in addition to removing litter. We measured plant productivity and thistle densities in frequently burned and unburned ungrazed tallgrass prairie. We found that the frequently burned sites were more able to resist invasion by the weedy species Cirsium arvense (Canada thistle) than the unburned site (P = 0.03). Within these two treatments, nitrogen manipulations appeared to influence thistle densities (P = 0.08). However, enhanced competition from tallgrass dominants under frequent burning appeared equally important.
Introduction
The grasslands surrounding the Boulder, Colorado area include all plant species associated with short-, mixed-, and tallgrass prairie (Bennett, 1997). Perhaps the most interesting of these grasslands is the tallgrass sites located in mesic bottomland areas. These sites possess the same dominant warm-season grasses as are found in mainland tallgrass prairie, and the sites are assumed to be relict grasslands that returned at the end of the Pleistocene (e.g., Hopkins-Arnold, 1998). Given the bottomland locations of these sites, these tallgrass areas are vulnerable to invasion by a variety of riparian woody species, and all grasslands in the Boulder area are threatened by a large number of nonindigenous plant species, many of which are considered noxious weeds (Bennett, 1997).
Fire is well known as an important component for maintaining grassland systems. Fire is also viewed by some researchers as a factor that can increase the likelihood of invasions by removing the plant canopy and creating a temporary fertilizing effect in the soil (Christensen and Burrows, 1986). This increased likelihood of invasions does occur under the circumstances of infrequent fire in an area where fire has been suppressed for a prolonged period of time. This response may not occur, however, in areas that are managed with a regular and frequent pattern of prescribed burns.
The responses to fire in areas where fire is infrequent and in areas where it is a regular occurrence are quite different. In a tallgrass prairie site where fire is infrequent, the fire suppression causes a build up of litter which lowers soil temperatures (Hulbert, 1988), reduces light levels near the soil surface (Schimel et al., 1991), and acts as a filter for nitrogen inputs from the atmosphere (Knapp and Seastedt, 1986). Grasslands which a have heavy litter accumulation tend to show a decline in grass species (Hobbs and Huenneke, 1992). In addition, areas with reduced fire frequency show increases in nitrogen availability in the soil, and increases in nitrogen levels in tallgrass prairie tend to cause increases in forb biomass (Seastedt et al., 1991).
In an infrequent burn, the fire will remove the litter layer on the soil surface and alter the microclimate and nutrient levels in the surface soil (Knapp and Seastedt, 1986). First, fire releases the light limitations created by the litter and, thereby, allows high-light, high nitrogen demanding species to more effectively compete. Second, it allows soil temperatures to increase, resulting in enhanced nitrogen mineralization from soil organic matter (Ojima et al., 1994). If there has been a large litter build-up due to fire suppression, the above two reactions will cause a "pulse" of nitrogen availability, which can allow invasive species to establish.
In contrast, frequent fire would not allow for the nitrogen build-up as nitrogen is not accumulated in the surface litter. Also, regular burning the plots is effective at increasing light intensity at soil level which causes increases in both vegetative and reproductive productivity of tallgrass species (Hulbert, 1988; Knapp et al., 1998). Increases in productivity of the tallgrass species will enable the plots to better withstand invasion by weedy species. Thus, we predict that regularly burned tallgrass prairie will be better at withstanding invasion than tallgrass prairie in which fire has been suppressed.
Methods
We measured Cirsium arvense L. (Scop) in 12, 5m x 5m ungrazed tallgrass plots that were established in 1993 (Hopkins-Arnold, 1998). Six of the plots were on the east side of the study site and the other six were on the west side of the study site in a slightly wetter area. Three plots on the east side and three on the west were burned four times over the course of the four years ('93, 95-97); the other three plots on each side were left unburned. Each plot on each side received one of the following six treatments from 1993-1997: burning alone, burning combined with addition of nitrogen fertilizer, burning combined with addition of carbon amendments, unburned plus nitrogen fertilizer, unburned plus carbon amendments, and control (unburned and no additions). Carbon amendments (sugar plus sawdust) act to reduce inorganic soil nitrogen levels in the soil (McLendon and Redente, 1992; Reever Morghan and Seastedt, in press). Nitrogen was added at 20 g N/m2 in 1993 and 1996 and 10 g N/m2 in 1994 and 1995. Sugar and sawdust were added in amounts that approximated a 25 g C/m2 per year addition in each of the four years (Hopkins-Arnold, 1998). The area surrounding the plots was burned in 1997, and this infrequent burn (the site had last been burned in 1993) was followed by a rapid increase in the density of Cirsium arvense throughout the study area.
To evaluate the effectiveness of the different treatments on resisting invasion, we compared forb and grass biomass amounts and densities of Cirsium arvense in response to the different treatments. The aboveground biomass in an 0.1 m2 section of each of the 12 plots was collected in 1997 and separated into grasses and forbs. In 1998 we conducted a census of Cirsium arvense in the twelve plots. We counted all Cirsium arvense in the full 5m by 5m plots and compared the density values for the different treatments to assess both fire and nitrogen status effects on the invasion.
Results
The biomass collections in 1997 indicated a much higher ratio of forbs to grasses in the unburned plots than in the burned plots (Fig. 1). There was a significantly higher biomass of forbs in unburned sites than in burned sites (P = 0.0125). There was also a significantly lower biomass of grasses in the unburned sites than in the burned sites (P = 0.0036).
The values for density of Cirsium arvense
were lowest in the plots which were burned and treated with carbon amendments
and were highest in the plots which were unburned and fertilized (Table 1). Regular burning correlated
more strongly with decreasing invasion than did decreasing the nitrogen
levels with carbon amendments. The decrease in density of Cirsium arvense
was significantly lower with regular burning (P = 0.03) but not quite significant
with manipulation of nitrogen via fertilizer and carbon amendments (P =
0.08). This trend was seen both on the east side and the west side, though
overall the plots on the wetter west side had higher densities of Cirsium
arvense than those on the drier east side.
Discussion
The results of this study support our hypothesis that regularly burned plots will be more resistant to invasion by Cirsium arvense than plots that were not regularly burned. The significant increase in forbs in the unburned plots (Fig. 1) supported previous research showing that fire suppression will cause a build up in nitrogen levels and an increase in forbs (Seastedt et al., 1991). The significantly lower grass biomass seen in the unburned plots (Fig. 1) supported statements of Hobbs and Huenneke (1992) that grasslands with a heavy accumulation of litter, such as the accumulation resulting from fire suppression, show a decline in grass species. The higher biomass of grasses in the regularly burned plots supports Hulbert's (1988) observation that production in tallgrass sites is higher in regularly burned areas. In our research area, the infrequently burned area has been burned twice to our knowledge since the establishment of the grazing exclosure in 1987, but likely was less frequently burned before that date.
The significantly lower densities of Cirsium arvense seen in the regularly burned plots as compared to the unburned plots (Table 1) also support our hypothesis by showing that the regularly burned plots were better able to withstand invasion by C. arvense. This ability to withstand invasion was very likely due to the combination of higher grass productivity and lower soil nitrogen availability in the regularly burned plots.
Subsurface irrigation appeared to play a role in facilitating invasion by Cirsium arvense. The plots in the wetter western side of the site had significantly higher thistle densities than those on the drier east side of the site. However, it is possible that the invasion began earlier at the west side of the plots and the thistle has had longer to become established. Another effect of subsurface irrigation may be higher production in this tallgrass site than would be expected by local precipitation levels. The average precipitation in this area is about 50 cm per year, and according to Sala et al. (1988) this amount of precipitation would correspond with tallgrass prairie production of about 400 g/m2. The mean production on these plots was 579.6 ▒ 74.1 g/m2, which may indicate some increase in productivity due to subsurface irrigation. Less water efficient forbs would benefit more from this enhanced water availability than more water efficient grasses. This irrigation water may also be a modest source of increased nitrogen.
Grassland sites are not all alike and not all will respond to regular burning in the same way. For example, a regular fire frequency in a site being invaded by a highly flammable exotic grass such as Bromus tectorum will only serve to favor the invader over the native grasses (D'Antonio and Vitousek, 1992). Yet in tallgrass prairie sites where the invaders are less fire tolerant than the natives, regular burning should prove effective at maintaining the native tallgrass species. We conclude that using fire properly in most tallgrass prairie sites will strengthen the ecosystem against invasion by an invasive species like Cirsium arvense rather than facilitating invasion.
Literature Cited
Bennett, B. 1997. Vegetation of the grasslands of the City of Boulder Open Space. Ph. D. Dissertation, Department of Environmental, Population and Organismic Biology, University of Colorado, Boulder. 128 p.
Christensen, P. E. and N. D. Burrows. 1986. Fire: an old tool with a new use, p. 57-66. In: R. H. Groves and J. J. Burdon (eds.). Ecology of biological invasions: an Australian perspective. Australian Academy of Science, Canberra, Australia.
D'Antonio, C. M. and P. M. Vitousek. 1992. Biological invasions by exotic grasses, the grass/fire cycle, and global change. Annual Review of Ecology and Systematics, 23: 63-87.
Hobbs, R. J. and L. F. Huenneke. 1992. Disturbance, diversity and invasion: implications for conservation. Conservation Biology, 6: 324-337.
Hopkins-Arnold, A. A. 1998. Response of Colorado tallgrass prairie to fire, winter grazing, and nitrogen manipulation. Ph. D. Dissertation, Department of Environmental, Population and Organismic Biology, University of Colorado, Boulder. 166 p.
Hulbert, L. C. 1988. Causes of fire effects in tallgrass prairie. Ecology, 69: 46-58.
Knapp, A. K., J. M. Briggs, J. M. Blair, and C. L. Turner. 1998. Patterns and controls of aboveground net primary production in tallgrass prairie, p. 193-221. In: A. K. Knapp, J. M. Briggs, D. C. Hartnett, and S. L. Collins (eds.). Grassland dynamics: long-term ecological research in tallgrass prairie. Oxford University Press, New York.
Knapp, A. K. and T. R Seastedt. 1986. Detritus accumulation limits productivity of a tallgrass prairie. BioScience, 36: 662-668.
McLendon, T. and E. F. Redente. 1992. Effects of nitrogen limitation on species replacement dynamics during early secondary succession on a semiarid sagebrush site. Oecologia, 91: 312-317.
Ojima, D. S., D. S. Schimel, W. J. Parton and C. E. Owensby. 1994. Long- and short-term effects of fire on nitrogen cycling in tallgrass prairie. Biogeochemistry, 24: 67-84.
Reever Morghan, K. J. and T. R. Seastedt. In press. Effects of Soil Nitrogen Reduction on Non-Native Plants in Disturbed Grasslands. Restoration Ecology.
Sala, O. E., W. J. Parton, L. A. Joyce, and W. K. Lauenroth. 1988. Primary production of the central grassland region of the United States. Ecology, 69: 40-45.
Schimel, D. C., T. G. F. Kittel, A. K. Knapp, T. R. Seastedt, W. J. Parton, and V. B. Brown. 1991. Physiological interactions along resource gradients in a tallgrass prairie. Ecology, 72: 672-684.
Seastedt, T. R., J. M. Briggs and D. J. Gibson. 1991. Controls of nitrogen limitation in tallgrass prairie. Oecologia, 87: 72-79.
Figure 1: Comparison of the ratio of forb biomass to grass biomass in the
unburned and regularly burned plots in tallgrass prairie at Boulder, Colorado.
| Āsource | Ādf | Āmean square | ĀPr > F | Ātreatment | Ān | Āmean ▒ se |
| ĀPlot | Ā1 | Ā57685.33 | Ā0.0066 | Āeast | Ā6 | Ā2.9 ▒ 1.0 |
| Ā | Ā | Ā | Ā | Āwest | 6Ā | Ā8.5 ▒ 1.9 |
| Ā | Ā | Ā | Ā | Ā | Ā | Ā |
| ĀBurn | Ā1 | Ā28812.00 | Ā0.03 | Āburned | Ā6 | Ā3.7 ▒ 2.1 |
| Ā | Ā | Ā | Ā | Āunburned | Ā6 | Ā7.7 ▒ 1.4 |
| Ā | Ā | Ā | Ā | Ā | Ā | Ā |
| ĀNitrogen | Ā2 | Ā14704.08 | Ā0.08 | Ā+ nitrogen | Ā4 | Ā8.1 ▒ 2.0 |
| Ā | Ā | Ā | Ā | Ācontrol | Ā4 | Ā5.7 ▒ 3.1 |
| Ā | Ā | Ā | Ā | Ā+ carbon | Ā4 | Ā3.3 ▒ 1.2 |
| Ā | Ā | Ā | Ā | Ā | Ā | Ā |
| ĀError | Ā7 | Ā3967.88 | Ā | Ā | Ā | Ā |
| Ā | Ā | Ā | Ā | Ā | Ā | Ā |
| ĀTotal degrees of freedom = 11 | Ā | Ā | Ā | Ā | ||
Table 1: ANOVA of treatment effects on density of Cirsium arvense. Mean values represent number of C. arvense individuals per square meter.
1 Current address: Graduate Group in Ecology, One
Shields Avenue, University of California, Davis, California 95616.
2 Current address: Graduate Program in Ecology, A118 Natural and
Environmental Sciences Building, Colorado State University, Fort Collins,
Colorado 80523-1499.
Back to the Home Page.