![]()
![]()
The gene curcuit method - model of the network of segmentation genes operating in the blastoderm of Drosophila
This model incorporates certain fundamental experimental observations
on the development of Drosophila melanogaster. Until the onset of
gastrulation, the Drosophila embryo has two unusual features. One
is the absence of cells: the syncytial character of the blastoderm permits
spatial interactions to be treated by the diffusive exchange of gene products,
and explicit cell- cell interactions are not present. The second is the
observation that although mutations of a segmentation genes alter the expression
patterns of other segmentation genes, they do not produce an alteration
in the morphology of the embryo until some time after gastrulation.
There are two hypotheses about the way that segmentation genes interact:
The authors considers the four gap genes Kr, kni, giant(gt) and hunchback
(hb) under the control of bcd and focus on that portion of the
blastoderm which gives rise to the segmented germ band. In this region,
the level of expression of gap genes is approximately a function only of
the position along the anterior-posterior (A-P) axis. Moreover the mutations
in the dorsal-ventral (D-V) genes do not affect the expression of the expression
of the A-P genes and vice versa. These observations suggest that the system
can be well represented by a linear array of nuclei running along the A-P
axis. The position of a nucleus is indexed by i.
During interphase the dynamical equation for ![]() the product in nucleus i, is given by
the product in nucleus i, is given by
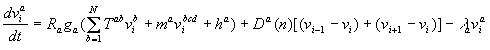
where N is the number of zygotic genes;
![]() is the matrix of genetic
regulatory coefficients; the elements of
is the matrix of genetic
regulatory coefficients; the elements of ![]() characterize the regulatory effect of gene b on gene a. This
matrix does not depend on i, a reflection of the fundamental fact
that the cell nuclei of a multicellular organism contain identical genetic
material.
characterize the regulatory effect of gene b on gene a. This
matrix does not depend on i, a reflection of the fundamental fact
that the cell nuclei of a multicellular organism contain identical genetic
material.
The bcd input is given by ![]()
![]() , where
, where ![]() is the concentration of Bicoid protein in nucleus i and
is the concentration of Bicoid protein in nucleus i and ![]() is the regulatory coefficient of bcd acting on zygotic gene a.
is the regulatory coefficient of bcd acting on zygotic gene a.
ga is a monotonic, saturating
function of the form 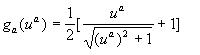 where
where
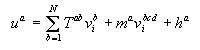
Ra is the maximum rate of synthesis
from gene a,
![]() is a threshold that controls
the constitutive activity of gene a.
is a threshold that controls
the constitutive activity of gene a.
The diffusion parameter ![]() depends on the number n of cell divisions that have taken place,
and varies inversely with the square of the distance between nuclei.
depends on the number n of cell divisions that have taken place,
and varies inversely with the square of the distance between nuclei. ![]() is the same for each species of protein, and invariant with respect to
n. It was assumed that the distance between adjacent nuclei is halved
after a nuclear division.
is the same for each species of protein, and invariant with respect to
n. It was assumed that the distance between adjacent nuclei is halved
after a nuclear division.
![]() is the decay rate of the
product of gene a.
is the decay rate of the
product of gene a.
The gene circuits method yields correct prediction of certain experimental results when applied to the problem of the formation of the segmental pattern in the fruit fly Drosophila.