![]()
![]()
This publication is devoted to one feature of the expression of Antp-like families of homeo box-containing genes (hox genes) in invertebrates and vertebrates and its consequences. An essential novelty of Lewis principle of collinearity of Antp-like family structure and pattern of their expression (Lewis 1978) is defined as following conclusion. A small but important part of the hereditary information is coded in consequence of hox genes which belong to the hox family (as in case of information coded in consequence of nucleotides in nucleic acids). This part of hereditary information describes a baseground of structure of developing organism and its metameric organization both in invertebrates and in vertebrates. Hence mutations in the hox loci lead to a cardinal break in basic structure of embryo body (Chisaka et all, 1991; Lufkin et all, 1991).
So far as concerns the genome of invertebrates, Antp-like families control the body plan along the only (antero-posterior) axis (Lewis 1978). The whole network of hox genes was formed from four Antp-like families on the macroevolutional way from invertebrates to vertebrates. Besides the organization of this HOX network is extremely conservative (Acampora et al. 1989).
We assume the transition from Antp-like family of insects to HOX network of vertebrates is connected with a macroevolutionary transition from the principle of collinearity to the principle of complanarity of the HOX network structure. This corresponds two-dimensional pattern of the hox genes expression in mesenchymal and neuroectodermal rudiments. The series of consequent duplications of initial ancestor's Antp-like family in vertebrate evolution conditioned qualitative complication of the feature of expression pattern of the HOX network from unidimensional (uniaxial) to two-dimensional (biaxial) "matrix" pattern. Thereby with each new duplication increase the number of lines in matrix of hox expression (from 2 to 4 and more). In other words, a well-defined topology of pattern expression of the hox genes follows from known scheme of the HOX network organization.
We suggest that biaxial, matrix organization of the HOX network pattern expression takes place in vertebrate development on repeated occasions: firstly in determination of axial mesenchyme and neuroectodermal structures and then in determination of limb buds and cranifacial structures.
In this paper we investigate the features of the suggested scheme of realization of information coded in structure of the HOX network on instance of mechanisms of determination of limb buds in mammals and birds in norm and experiments.
The computer program of HOX network behavior in the mesenchyme derivatives of vertebrate embryos describes following levels of biochemical signaling in the processes of hox genes expression:
As complicated system with features of synergetic system and cybernetic (boolean) network the modeled hox network shows several modes of the patterning processes. It may be correspondence between founded steady states and diverge spatio-temporal patterns of real hox network expression in vertebrate embryogenesis.
Chicken wing development in norm: 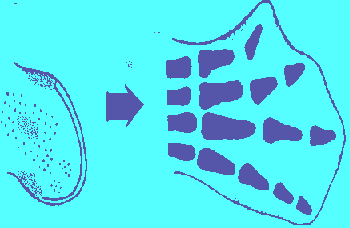
Presumptive Map for chicken limb bud: 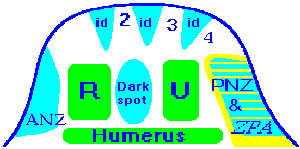
We consider the possibility that the spatial pattern of expression of the HOX families in embryogenesis of mammals and birds has the form and function of coordinate grid. Function of progress zone of the limb bud is the production of cell material with "striped" pattern of determination. So that along proximo-distal axis each Hox family is expressed in its own zone (stripe). In present case the stripe of expression of Hox-3 family corresponds to presumptive zone of shoulder, Hox-4 - presumptive humerus, ulna and radius, Hox-1 - presumptive wrist elements, Hox-2 - digit-forming region.
On the other hand, function of polarizing region is initiation of consequent
activation of hox genes along anteroposterior axis. The results of computer
reproduction of events of hox patterning lead to principal conclusion that
each domain of the hox gene expression marks either zone of condensation
of cartilaginous elements or necrotic zone. Particularly, hox-3.3 + -3.1
gene expression region marks coracoid (CO) determination
zone; hox-4.9 marks proximal epiphysis of humerus; hox-4.1 - diaphysys
of humerus; hox-4.2 - distal epiphysis; hox-4.6 region of expression corresponds
to zone determination of ulnae (U); hox-4.4 - zone
determination of radius (R). Nine genes in Hox 2 family
corresponds nine zones of determination in digit forming region of mammals
limb buds (five fingers 1 2 3 4 5 and
four interdigital necrotic zones). In the case of wing bud and corresponding
wing presumptive regions predicted Hox pattern is presented on Fig.1. 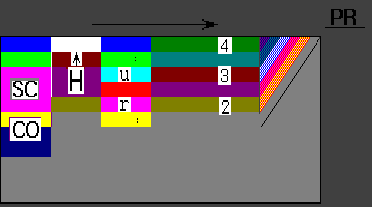
Normal Chicken wing skeleton as is the case of Fig.1: 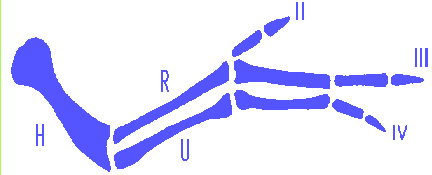
Apparently the hox pattern of chicken limb buds differs from mammalian one by reduction and elimination of territories of expression of 5' and 3' marginal hox genes, which determine digits 1 and 5, posterior necrotic zone and fibula.
A wide variety of abnormalities in the pentadactyl limb skeleton (conditioned by mutant genes, chemical agents or surgical manipulations) are enable to verify the predictions of the model. Changes in the time interval of competence of limb mesenchyme to induction of hox gene expression or the shift of polarizing region (PR) along competent tissue or splitting of the PR or local breaking in mechanisms of the competence time determination all permit to explain the variety of abnormalities in the limb bud morphogenesis.
As example we consider the results of computer simulation of Wolpert and Hornbruch experiments (1981; 1987). These authors examined the effect of grafting a wing PR to different position along anteroposterior axis of the limb bud.
The humerus (H) between the host (PR)
and grafted polarizing region (PR') often showed mirror-image
symmetry along the anteroposterior axis. The radius was absent in most
cases, the ulna (U) was usually duplicated and the
pattern of digits was often 4 3 3 4
. The result of our computer simulation is in close agreement with that
experiments (Fig.2). 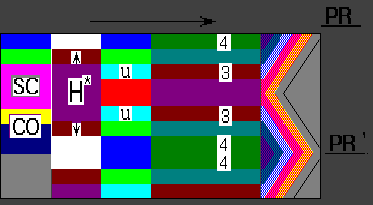
The Map of the limb bud determination shown in Fig.2 corresponds "monster's"
wing similar with such: 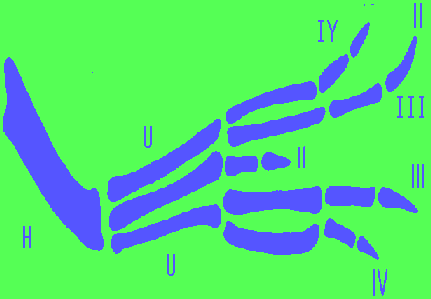
The limb that developed following the grafting of polarizing regions
(PR' and PR") opposite somites
16 and 17 leads
to the pattern of digits 4 3 4 4 3 3 4 ,
duplication of ulnae and absence of radius (Wolpert and Hornbruch, 1981).
It is in good agreement with the simulation results (Fig.3). 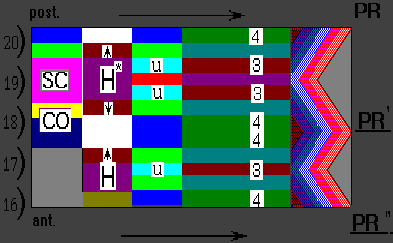
The Map of the limb bud determination shown in Fig.3 corresponds "monster's"
wing like this: 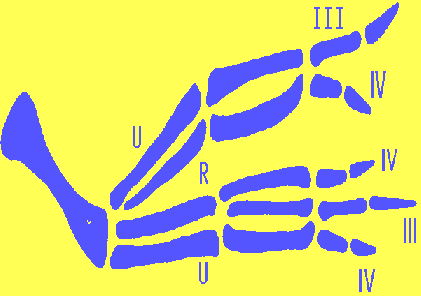
The model describes other effects of PR-grafting to different positions along the chicken limb bud and effects of multiple grafting of PR on the digit patterning in experiments of Wolpert and Hornbruch (1981; 1987). Analogously it simulates RA-induced coracoid duplications and "homeosis" transformation of coracoid into the scapula (mirror-image transformation) in experiments of Oliver et all (1989). The model predicts the mechanisms of the majority of polydactylous mutations in human, rodents and fowls (such as pre- and postaxial polydactyly, fowl talpid polydactyly; guinea-pig "polydactylous monster"), as well as preaxial poly- and ectrodactyly coupled with preaxial hemimelia being result of treatment of chemical teratogenic agents.
There are the reasons for application of the hox gene grid conception to another patterning processes in vertebrate development.
![]()
![]()