Los canales iónicos son proteínas integrales de la membrana que permite el paso selectivo de algunos iones, en la presencia de algún estímulo. Son principalmente importantes en células excitables, como las neuronas y los músculos. De hecho, todo el trabajo de las neuronas está relacionado a la función de los canales iónicos. Los canales iónicos catalizan directamente el flujo de iones que provoca los cambios de voltaje a través de una membrana y, simultáneamente actúan como sensores de señales fisiológicas, como cambio de voltaje o cambio en la concentración de un ligando, organizando los cambios eléctricos de la neurona. (Miller, 1992)
Mientras que la célula
gasta una gran parte de su energía para mantener los
gradientes de concentración de iones, son los canales iónicos
quienes se gastan esa energía, al abrirse y permitir el flujo
de iones. (Clapham, 2000) Los canales pueden ser fácilmente
comparados con enzimas, ya que un pequeño cambio en la
conformación de su "puerta" puede abrirla o
cerrarla, llegando a tener una eficiencia de hasta
![]() iones / segundo (Clapham, 2000) La eficiencia de los canales iónicos
es tan grande, que generalmente una célula tan solo necesita
un par de miles de ellos, contrario a una enzima, de la cual se
necesitan concentraciones de varios órdenes de magnitud.
iones / segundo (Clapham, 2000) La eficiencia de los canales iónicos
es tan grande, que generalmente una célula tan solo necesita
un par de miles de ellos, contrario a una enzima, de la cual se
necesitan concentraciones de varios órdenes de magnitud.
Actualmente se conocen dos tipos principales de canales iónicos. Los primeros son los operados por voltaje, y los otros los operados por ligando. (Miller, 1992) De los últimos, todavía se diferencian los activados por un segundo mensajero intracelular y por un ligando externo. (Clapham, 2000) . Lo que se conoce acerca de ellos todavía es muy limitado. No existe ninguna imagen de alta resolución de estas proteínas, así que casi todo lo que se sabe se basa en pruebas fisiológicas, que han permitido crear un modelo estructural de los canales. Se cree que son más o menos agregados cilíndricos con varias subunidades, con un poro de transmisión iónica en el eje de simetría. (Miller,1992)
Un buen ejemplo que ilustra la estructura de la mayor parte de canales es el canal de potasio activado por voltaje. Este cuenta con 4 unidades alpha, cada uno con 6 regiones hidrofílicas (S1-S6) y 4 subunidades Beta hidrofóbicas. Entre el S5 y S6 existe un segmento H5. (Yeh Jan, 1994)
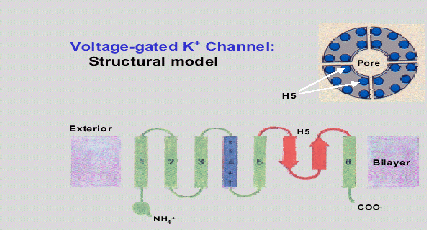
Se sabe que los residuos en cada subunidad, en posición equivalente tienen una influencia electrostática similar, que favorece el paso del ion. Adicionalmente, se sabe que son proteínas transmembrana, que la suelen atravesar varias veces, aunque es sólo por un segmento que sucede el transporte de iones.
Los iones se comportan como cualquier substancia: tratan de irse de un lugar de mayor a otro de menor concentración. Sin embargo, por ser moléculas cargadas, también son influenciados por campos eléctricos. Cuando las fuerzas provocadas por el gradiente químico y el campo eléctrico se equilibran, se dice que se llega al equilibrio de Nernst. Debido a que cada ion tiene un potencial de Nernst distinto, el hecho que se abra un canal va a cambiar todo el potencial de la membrana de la célula. (Clapham, 2000) Los potenciales de Nernst se puede calcular con la siguiente fórmula:
 ,
donde Z es la carga del ion, [C1] la concentración interna, y
[C2] la externa.
,
donde Z es la carga del ion, [C1] la concentración interna, y
[C2] la externa.
Como cualquier enzima, los canales iónicos no alteran el flujo natural de los iones. Sin embargo, permiten que este suceda a través de una membrana que lo impide, y además hace que ocurra rápido. Sin embargo, el canal iónico tiene un mecanismo para favorecer el flujo de un lado hacia el otro. (Yeh Jan, 1994) Esto no significa que fuerce a los iones a ir en contra de su gradiente. Simplemente, tiene una estructura tal que impide en cierta medida el paso de iones en la dirección no deseada. Este sistema está relacionado a la forma en que se cierran las compuertas cuando ya no es deseable que siga abierto. Sin embargo, mientras el canal está abierto, existe flujo de iones bidireccionalmente, siendo obviamente, el flujo neto, según el gradiente electroquímico del ion.
La activación de la compuerta de un canal puede ser compleja. Se cree, que para canales operados por voltaje, hay un cambio conformacional que tiene como última consecuencia el paso del ion. Primero, el segmento S4 actúa como un dipolo eléctrico. Bajo cierto voltaje existe un momento de dipolo lo suficientemente grande para causar un cambio de conformación. El residuo cargado de esta proteína cargada se mueve a través de la membrana, cambiando la estructura terciaria del canal. Este cambio permite que un ion (el canal es selectivo) se mueva interactuando con las otras subunidades del canal, hasta atravesar la membrana en su totalidad. (Stemmler, 1998) El tipo de ion que transporta la membrana es decidido por la posición que ocupa el residuo responsable del dipolo en S4.
Como ya se dijo, el canal es muy selectivo. El canal de potasio operado por voltaje es al menos 100 veces más permeable al iones de potasio que a los de sodio. (Yeh Jan,1994) Debido a que el radio del ion de sodio es más pequeño que el de potasio, es difícil pensar que la selectividad se deba al "tamaño" del conducto. Se cree que el canal tiene múltiples puntos de adhesión a través de su largo transmembrana. (Yeh Jan,1994) Se puede asumir que la velocidad de paso de iones depende únicamente de la concentración de iones de potasio. Sin embargo, esto no es cierto. La presencia de otros iones tiene influencia en la velocidad de paso. (Yeh Jan,1994) Si existe otro ion que es permeable, pero que tiene mayor afinidad a los puntos de adhesión, esta especie va a tardar más tiempo adentro del canal, bajando la velocidad de todo el canal. Contrariamente, si sólo hay iones potasio, estos van a adherirse a los puntos de adhesión, y luego van a causar repulsión contra otros potasio, obligándolos a pasar más rápido.(Yeh Jan,1994) Es importante notar que la concentración de potasio necesaria para que se llenen todos los puntos de adhesión del canal es relativamente baja, lo que hace que trabaje casi todo el tiempo a su velocidad máxima y que aparente se constante.
En un sistema, la velocidad del paso de iones es proporcional al número de canales abiertos en toda la membrana celular. El número de canales abiertos, está determinado por una ecuación diferencial de primer orden. Este modelo, es válido sólo en canales donde existe una conformación abierta, y otra cerrada, y donde el estado de energía libre de la proteína es el mismo, tanto para la conformación abierta como para la cerrada. (líneas punteadas en la gráfica) (Stemmler, 1998) Existen un gran número de fórmulas para determinar la probabilidad que un canal se abra, y estimar bajo cierto voltaje el número de canales en la célula que se abrirían.

Una vez se abre el canal, debe existir un mecanismo para cerrarlo. El mecanismo más aceptado es el de la cadena y la bola. (Yeh Jan,1994). Según este modelo, una compuerta, del lado citoplasmático del canal se cierra. Esta compuerta suelen ser los primeros 20 residuos de una subunidad alpha o beta (o las dos). Un canal iónico se puede volver a abrir, por lo que el canal, al cerrarse, no forma ningún enlace covalente nuevo. Como se mencionó anteriormente, el sistema de cerrar las compuertas está relacionado con el desfavorecer el flujo del ion en cierta dirección. Cuando la concentración del ion cambia, también ocurre un cambio en el potencial de Nernst para este ion. Esto causa el cambio conformacional ya descrito. (Yeh Jan,1994) Sin embargo, al menos en los canales de potasio operados por voltaje, este no es el único mecanismo para cerrar el canal. El otro mecanismo parece no ser tan usado por la célula. Este se basa en un ion de magnesio que tapa el canal. El magnesio se encuentra en el citoplasma, y en ciertas condiciones se ve atraído al canal. Sin embargo, este no puede pasar, teniendo como resultado, cerrar la compuerta. (Yeh Jan,1994)
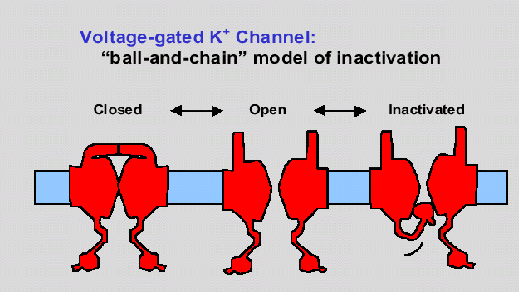
Hasta el momento, solo se han ilustrado ejemplos con canales operados por voltaje. Sin embargo, se sabe que en los canales operados, la estructura es similar. La diferencia radica en que en vez de ser un cambio de potencial, lo que activa al canal es la presencia de una molécula. Esta molécula causa un cambio electroquímico, que sería equivalente al cambio conformacional realizado por el dipolo. El ejemplo de canal operado por ligando más estudiado es el de la acetilcolina. Este canal se encuentra en las células excitables -- músculos y neuronas.
El receptor de acetilcolina consta de 4 subunidades transmembrana, dispuestas en forma de pétalo, con un centro, en el cual se cree que pasa el ion de sodio cuando se activa. (Changeux, 1993) Adicionalmente, en uno de las otras subunidades, está el receptor de la acetilcolina. Las cuatro unidades, hidrofílicas se les conoce como S1-S4. El receptor de acetilcolina se encuentra en dos de estas unidades, y es necesaria la presencia de dos acetilcolinas para que se abra el canal. (Changeux, 1993) Además de las subunidades transmembrana hidrofílicas, tiene 4 segmentos hidrófobos, que rodean a los hidrofílicos para "protegerlos" del lípido de la membrana. Estos segmentos se les denominan M1-M4. El canal está regulado por los cambios en la posición de estos segmentos.
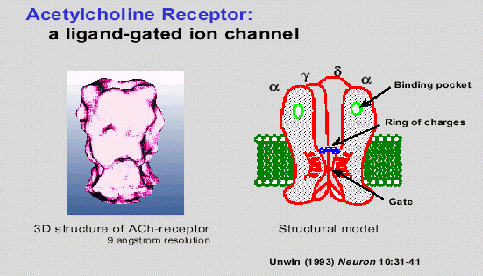
Los receptores de acetilcolina presentan varios cambios conformacionales. Además de sus estados abierto-cerrado, tiene un estado cerrado-saturado. Cuando la concentración de acetilcolina es constante durante un largo período de tiempo, la proteína se protege, y evita que continúe el paso de iones. (Changeux, 1993) Obviamente, este tercer estado introduce complicaciones a la cinética de la proteína. Según Changeux y colaboradores, este tercer cambio puede estar involucrado en el aprendizaje. Si bien, no se han presentado pruebas convincentes que esto sea así, tampoco se ha podido probar lo contrario. (Changeux, 1993)

La regulación de los canales iónicos también puede estar regulada por la fosforilación de la proteína. Existen algunos canales iónicos que cuando son fosforilados dejan de funcionar. (Changeux, 1993)
En resumen, la mayor parte de canales iónicos presentan una estructura similar, que consta de varias subunidades transmembrana que forman un canal a través del cual pasa el ion. Un agente externo activa el canal, el cual deja pasar selectivamente a un ion. Después de cierto tiempo o estímulo, el canal se cierra de nuevo, mediante otro cambio en su conformación.
En los vertebrados, los canales iónicos se encuentran en prácticamente todas las células. Hay en las neuronas, otros regulan el latido cardíaco, las funciones de los túbulos renales, entre otros. (Changeux, 1993) Obviamente, cualquier problema en la función de los canales iónicos puede tener consecuencias desastrosas. Entre esas enfermeades, están:
miotonia: el músculo tarda mucho tiempo en relajarse después de la contracción. Es causado por problemas en el transportador de cloro o de sodio en las células musculares.
Fibrosis cística: Problemas en el canal de cloro causan acumulación excesiva de agua en el tracto respiratorio, propiciando infecciones.
Síndrome de Prader-Willi: Canales de cloro defectuoso causan condiciones neurológicas severas. (Canales de cloro activados por GABA)
Hipertermia maligna: Canales de calcio en el músculo no funcionan bien. Se eleva la temperatura exageradamente, principalmente durante una anestesia.
(Neuromuscular Disease Center, 2001)
Referencias
Changeux, Jean Pierre. 1993. Chemical Signaling in the Brain
Sci. American Nov. 1993 30- 37
Clapham. 2000 Background: Ion Channels
Harvard University
http://clapham.tch.harvard.edu/ionchannels.htm
Lilly Yeh Jan, Yuh Nung Jan. 1994. Potassium channels and their evolving gates
Nature 258:119-122
Miller, Christopherr. 1992. Ion Channel Structure and Function
Science 258:240-241
Neuromuscular Disease Center. 2000. Ion Channel Diseases
Washington University School of Medicine
http://www.neuro.wustl.edu/neuromuscular/mother/chan.html
Stemmler, Martin. 1998. Kinetics of Two-State Ion Channels
California Institute of Technology
http://www.klab.caltech.edu/~stemmler/s4node2.html