| THE BASIC NEURON |
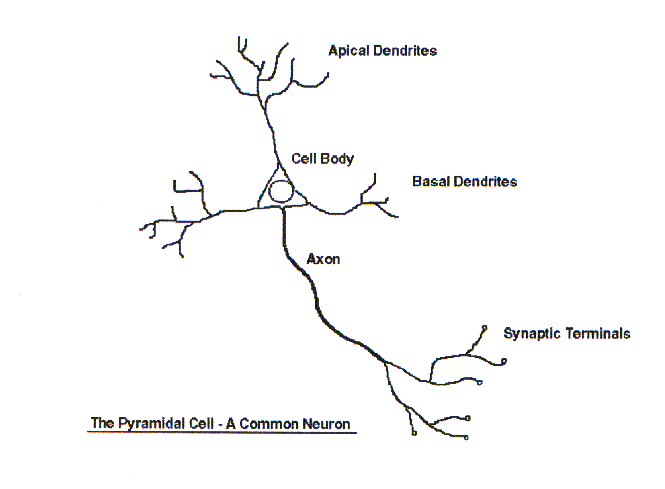
Neurons are the primary cell that involved in information processing in the brain. An important quality of neurons is that they are not replaced by new neurons if they are damaged - brain function is repaired by way of using existing cells. There are many different types/shapes of neuron, but the basic structure is always the same. Figure 1 shows a pyramidal cell, a common neuron in the class of cortical neurons. The neuron is mostly made up of components which only vary in their size and number between different types.
 The Cell Body
The Cell Body
The cell body contains the biochemistry that makes the cell operate. It varies in size and shape, depending on the functional operation of the cell.
The Dendrites
Dendrites are the 'input lines' of a neuron, they take signals from sensory input, or other nerve cells, and integrate then. A nerve cell can have many dendrites which branch many times, their surface is irregular and covered in dendritic spines which are where the synaptic input connections are made.
The Axon
The axon is similar in shape to dendrites, but is the 'output line' of the neuron, and it is smoother on its surface, longer, some stretching many feet, and there is only one on each cell. It usually only starts to branch off towards its end, and there are fewer branches.
The Axon Hillock
The axon hillock is where the axon is joined to the cell. It is from here that the electrical firing known as an action potential usually occurs.
The Synapse
Synapses are the junctions formed with other nerve cells where the presynaptic terminal of one cell comes into 'contact' with the postsynaptic membrane of another. It is at these junctions that neurons are excited, inhibited, or modulated. There are two types of synapse, electrical and chemical.
Electrical synapses occur where the presynaptic terminal is in electrical continuity with the postsynaptic. Ions and small molecules passing through thus connecting channels from one cell to the next, so electrical changes in one cell are transmitted almost instantaneously to the next. Ions can generally flow both ways at these junctions i.e. they tend to be bi-directional, although there are electrical junctions where the ions can only flow one way, these are know as rectifying junctions. Rectifying junctions are used to synchronise the firing of nerve cells.
The chemical synapticjunction is more complicated. The gap between the post- and presynaptic terminals is larger, and the mode of transmission is not electrical, but carried by neurotransmitters, neuroactive substances released at the presynaptic side of the junction. There are two types of chemical junctions. Type I is an excitatory synapse, generally found on dendrites, type II is an inhibitory synapse, generally found on cell bodies. Different substances are released at these two types of synapse. The direction of flow of information is usually one way at these junctions.
Two Types Of Neuron
There are two main types of neuron: Golgi Type I Neurons and Golgi Type II, also called association neurons. The Golgi Type I neurons are long-axon neurons which carry information from one part of the brain to another, or from the brain, or spinal cord to effector organs such as muscles. They are larger than Type II which have short, sometimes no axons, which are used locally for processing and integrating information. There are many subtypes of neuron which fall under these two categories. Brains of more highly developed animals contain more of Type II than Type I cells. The processes of Type II cells are contained in the nucleus and the terminals receive and make synaptic contacts. On Type I, synaptic contacts are only generally found on dendrites and the cell body. In Type I neurons, the dendrites only receive, the axons only transmit, in type II neurons all connections with other cells tend to both transmit and receive, i.e. they can be post- and presynaptic. A result of this characteristic is that interaction with another cell can be handled by one dendrite, or part of the axon terminal without the need for the entire cell to be used.
ELECTRICAL ACTIVITY IN NEURONS

Figure 2 - Electrical Exchanges In The Nervous System - Source
Information is carried and encoded in neurons by the use of electrical signals.
Underneath the generation of these signals is a voltage maintained in the cells themselves, the inside of a cells is negative to its outside when it is at rest, this voltage is known as the resting potential (-70mV). There are other kinds of potential involved in transmitting these signals. Figure 2 shows the electrical activity involved in the function of the nervous system. Sensory stimuli act on receptors, which reply by creating receptor potentials, these in their turn cause what is known as action potentials which are what carry information larger distances through the nervous system. As information passes from one neuron to another at synaptic junctions, synaptic potentials are caused in postsynaptic neurons and cause action potentials in postsynaptic cells. The receptor potentials can also directly evoke synaptic potentials in adjacent neurons, and also where input and output synapse distances are not great, the synaptic potentials themselves can generate others of a similar nature. These synaptic potentials can lead to activation of effectors such as muscles, resulting in behaviour. Although receptor and synaptic (also known as generator) potentials and action potentials are both electrical in nature, they have very different behaviours.
Differences Between Generator Potentials and Action Potentials
Generator Potentials:
Amplitudes are graded with stimulus intensity; the stronger the stimulus, the larger the potential. All potentials are the same size, regardless of stimulus strength. Potentials may last for as long as the stimulus lasts. Potentials are of constant duration. Tiny potentials may be evoked with the smallest possible stimulus.
Action Potentials:
Action potentials require a substantial change in membrane potential for their generation. Potentials sum when two stimuli are presented together. A refractory period of about 1.5 ms occurs after every action potential during which time another action potential cannot be generated, regardless of stimulus length. Potentials spread passively from the site of generation; they are the largest where they are produced and become progressively smaller away from that point. Potentials are actively regenerated along an axon; a potential recorded at the end of an axon is the same size as one recorded at the beginning of an axon.
Membranes and Membrane Channels
All electrical potentials in biological systems are generated across cell membranes. The potentials are created by the movement of ions across membranes, this is done through channels. There are five basic types of membrane channel:
Leak channels account for the natural permeability of membranes to ions. Voltage sensitive channels vary in permeability depending on membrane voltage, and can be found mainly on axons. Ligand-sensitive channels respond to specific chemical agents and open or close in the presence of the agent, and can be found on dendrites and cell bodies at postsynaptic sites. Mechanosensitive channels respond to deformation of the channel or membrane surrounding it, and are found in receptor cells. Gap-junctional channels allow ions to move between cells, especially at electrical synapses.
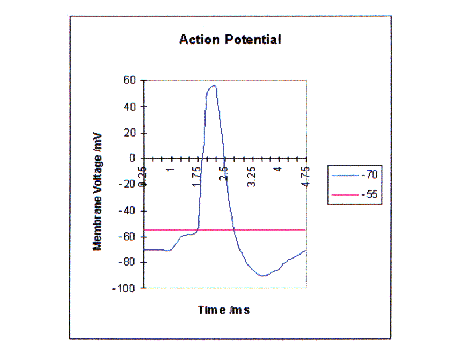
Figure 3 - Voltage Changes Involved In An Action Potential
Action potentials are transient, all-or-none voltage impulses that neurons employ to transmit information down an axon.
When a small amount of current is passed into an axon, so that the change induced in the membrane potential is less than 15mV, the response is passive - the voltage change reflects the resistive and capacitive properties of the membrane. However, if the voltage is changed by more than 15mV in the positive (depolarising) direction then an explosive change occurs in the potential of the cell. Negative current do not evoke such a change.
The resting potential is -70mV, if this is changed by more than 15mV, i.e. the potential in the cell goes above -55mV, the change in potential becomes rapid. The cell membrane voltage becomes rapidly positive, going up to 50mV in about 1ms and falling again, it then falls even further than the resting potential, to -90mV in another millisecond and only then returns to the resting potential. This massive voltage change is called the action potential.
This voltage change is due to voltage-dependant permeability changes in the cell membrane. A variety of both positively and negatively charged ions are present in both the cell and its surroundings. Na+, sodium, K+ potassium and Cl- chlorine being the most important. Nerve cells actively maintain a high concentration of K+ ions inside the cell and low concentration outside, so K+ ions tend to flow out of the cell. With depolarisation, the membrane conductance to Na+ ions increases. With the increase in Na+ conductance, Na+ ions enter the axon and cause more depolarisation; this leads to increasing the Na+ conductance even more, a positive feedback loop which results in the accelerating, regenerative response. However it is when the current coming in as a result of the Na+ ions exceeds the rate at which K+ ions are leaving the cell that the response becomes regenerative, i.e. when the axon is depolarised by 15mV, the threshold and then the action potential explosion occurs. Following the initiation of the action potential, the membrane permeability to Na+ ions remains high for only a short time, after which it then becomes more permeable to the K+ ions and so the membrane voltage falls . As the membrane voltage depolarises, the increased K+ conductance decreases and the resting potential of -70mV is regained. Both Na+ and K+ conductances are dependant on both voltage and time. Cl- conductance does not change and plays no role in generating action potentials. The action potential is usually generated at the axon hillock.
Synaptic Potentials
As mentioned before there are two types of synaptic junction, inhibitory and excitatory: a small depolarisation occurs at membrane sites postsynaptic to an excitatory synapse, called EPSP (excitatory postsynaptic potential); at sites postsynaptic to inhibitory synapses, the effect is to hyperpolarise IPSP (inhibitory postsynaptic potential). Individual EPSP's and IPSP's are two small to have much effect on a neuron, but when they are summed, they change the postsynaptic membrane potential substantially. Therefore the output of the cell reflects the balance of all the inputs, inhibitory and excitatory, the first trying to lower membrane potential, the latter to raise it. The EPSP's are dependant on the Na+ and K+ ions, IPSP's are dependant on the Cl- ions.
At electrical synapses, current moves relatively unimpeded across gap junctions. Gap-junction channels are the means by which the current flows, and they can stay open for long periods of time, up to 100ms. It is often impossible at these junctions to distinguish the post- and pre-synaptic sides, both sides of the junction can be either post- or presynaptic and current can flow both ways.
| CHEMICAL ACTIVITY IN NEURONS |
Synaptic Transmission Chemistry |
Most synapses in the brain are chemical. A substance is released at a synapse and diffuses across and instigates a change in the postsynaptic membrane. Some substances act directly on postsynpatic membranes, altering the permeability and allowing ions to flow - these are known as neurotransmitters. These are what control the EPSP's and IPSP's and they do not generally last much longer than 10- 100 ms. Other substances modify neural activity rather than initiating it. These are called neuromodulators and they can last for minutes, hours, or longer; their onset is slow, a matter of seconds. Biochemical changes are induced in the neurons that alter membrane channels, enzymatic activity and perhaps gene transcription. Neuromodulatory activity has good evidence to suggest that it underlies memory and learning.
Most animals have a number of different sensory receptors each responding to various stimuli - light, sound, touch, heat. These receptors transduce the stimulus into a sustained, graded receptor potential. All neurons have characteristics of sensory receptors in that they have specialised areas of membrane which respond selectively to specific channels, neurotransmitters and neuromodulators. The different types of sensory receptors respond in different ways, e.g. the touch and auditory receptors respond in a similar way to the neurotransmitters affects on neurons, whereas the photo- and olfactory receptors react to stimuli like neurons to neuromodulators.
These include touch, pressure and auditory receptors. Contained within a the membrane of a bare nerve ending of a cell of this type are specialised channels whose conductance is altered when they, or the surrounding membrane are deformed. A rest, few if any of the channels are open, but when the membrane is stretched the channels change configuration and allow ions to flow. Voltage sensitive channels are located on the cell also, and when sufficient depolarisation is generated, actions potentials arise. Na+ and K+ are involved in this process.
Neurotransmitters and Neuromodulators
Receptor Potentials
Mechanoreceptors
|
Photoreceptors |
Invertebrate and Vertebrate Photoreceptorsdiffer dramatically in their operation.
Invertebrate photoreceptors respond to light. Visual pigment molecules found in specialised membrane absorb light and lead to an influx of Na+ ions into the receptor cell. In weakly-illuminated, dark-adapted receptors, small depolarising responses to single quanta are detected. In brighter light, the quantal responses, bumps, summate, causing the receptor potentials. The quantal bumps of dark- adapted cells are larger than those of receptors just exposed to light, and hence explain the decline of receptor potential in the presence of continuous light.
Vertebrate photoreceptors are hyperpolarised by light and are depolarised in the dark. Channels in the cell's membrane remain open in the dark, admitting Na+. Current flows into the vertebrate receptor in the dark and the dark current is turned off by light.
Adaptation is important in all receptors. With a sustained stimulus, the receptor potential declines. There are three types of adapting receptor, fast, medium and slow.
Adaptation
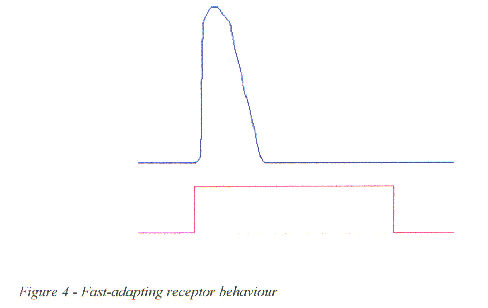
Figure 4 - Fast-adapting receptor behaviour
Fast-adapting, or phasic receptors respond only to a change in stimulus level, as are most touch and olfactory receptors, i.e. the receptor responds immediately on application of the stimulus, but rapidly decays to zero even though the stimulus is maintained.

Figure 5 - Medium-adapting receptor behaviour
Medium-adapting receptors decline much more gradually in the presence of a long stimulus, it can decay to zero if the application is sufficiently long. Taste and hearing receptors are like this.
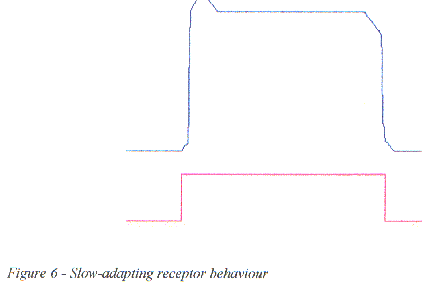
Figure 6 - Slow-adapting receptor behaviour
Slow-adapting, or tonic receptors, such as photoreceptors respond to a prolonged stimulus in two stages, an initial transient potential that decays to a smaller plateau potential which is maintained for as long as the stimulus lasts. All receptors respond maximally to changes in stimulus level.
created by Diana Weedman Molavi, PhD

