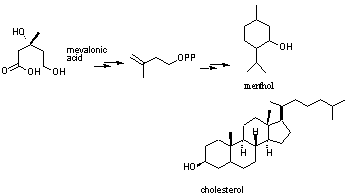
agents and pheromones.Classically terpenes are biosynthesised from mevalonic acid through to IPP which then repeatedly
combine.
2 Incorporation of substrates - determination of which pathway is operative in each system
3 Feeding of putative intermediates
4 Conclusions
Elucidation of biosynthetic
pathways - the new terpene pathway
| Terpenes are a diverse class of secondary metabolites numbering over
30,000 that have been known since ancient times. Terpenes are widely distributed
throughout Nature occurring in microbial, plant and animal systems.Inplants
they are represented as essential oils whilst in animals terpenes are processed
on to form steroids. Many terpenoid substances are physiologically active
such vitamins A, K and Q. Biologically they are used as antifeedants, defensive
agents and pheromones.Classically terpenes are biosynthesised from mevalonic acid through to IPP which then repeatedly combine. |
|
The new biosynthetic pathway
to terpenes
Discovery of a mevalonate
independent pathway
|
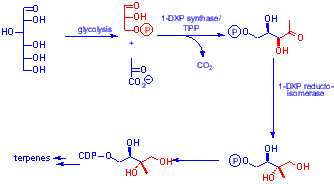
It has recently been shown that some isoprenoids are biosynthesised through 1-D-deoxy xylulose-5-phosphate (DXP) rather than mevalonic acid1. DXP is formed from the thiamine pyrophosphate mediated decaroxylativeaddition of pyruvate to glyceraldehyde-3-phosphate. Methyl erythritol-5-phosphate (MEP) has been shown to be the next intermediate on the pathway after DXP -the product of the action of the enzyme DXP reductoisomerase2 . |
 |
Clues to the pathway
Feeding experiments performed
| The steps converting MEP to IPP are unknown
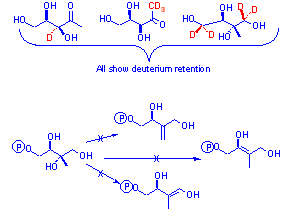
although the deoxygenative processes cannot involve dehydration as [2H] labelled deoxy xyluloses and methyl erythritols fed to bacteria show 2H retention3. As the deuterium atoms are
|
 |
Our systems
The systems used and the metabolites
produced
|
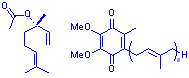
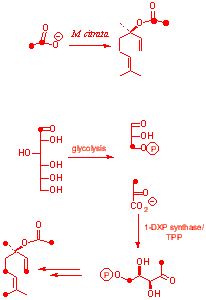
We use plant tissue cultures of Mentha citrata transformed with the bacteria Agrobactrium tumefascians. After approximately 25 days of growth approximately 6mg of the mixed metabolite linalyl acetate is produced and extracted. E. coli is also used and produces ubiquinone- 8 which is also known as vitamin Q. We also use |
 |
MY RESEARCH
Aims of the research
Establish which pathway is operative in bacterial,
plant and fungal system
Determine nature of post MEP intermediates using
feeding experiments with 13C and 2H labelled compounds
Feeding of [1,2-13C2] acetate and [1-13C]
glucose
Determining which pathway is operative
in M. citrata
| Feeding of 13C2 sodium acetate to M. citrata
showed incorporation into the acetate unit only.
[1-13C] glucose showed incorporation of 13C
|
 |
Features of the pathway in plants I
Using labelled substrates to investigate
biosynthetic steps
|
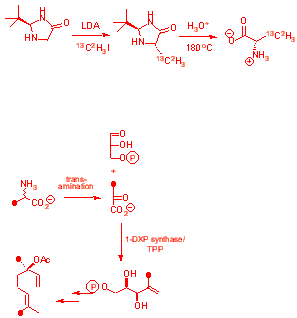
As for bacteria, MEP must be deoxygenated, but the mechanism of how was unknown. Alanine was used as a pyruvate surrogate to probe the C1 methyl group of DXP. [3-13C] alanine shows processing to pyruvate
[3-2H3] and [3- 13C2H3] alanine show retention
|
 |
Features of the pathway in plants II
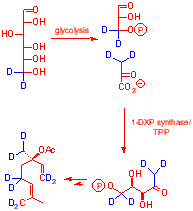
| [6,6-2H2] glucose labels the protons of
C5
of DXP via glycolysis and was used to determine whether the C5 protons of DXP are abstracted in post MEP processing GC-MS showed a high (5.96%) M+2 ion,
|
 |
Results of feeding intermdiates
The intermediates were synthesised by Dr Andy Humphrey and I
and fed to both systems
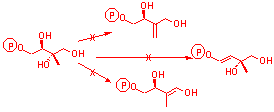
| Known Intermediate on mevalonate independent pathway | Intermediates common to both pathways as pyrophosphates | Putative intermediates on the mevalonate independent pathway |
![]()
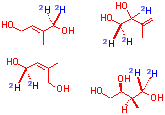
E. Coli
Incorporation
M. Citrata
Incorporation into neither E Coli nor M. citrata
No Incorporation
The lack of incorporation of the free alcohols of IPP and DMAPP which are known to be intermediates on the pathway show that both E.coli and M. citrata lack the necessary kinase to phosphorylate later intermediates fed as alcohols. Given this, it is not possible to rule out any of the above as potential intermediates.
Conclusions about the new pathway in
plants
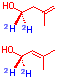
| Retention of deuteruim atoms
from labeled glucose and alanines show the deoxygenative step from MEP to IPP cannot proceedvia dehydrations in plants. Thisplaces limitations on the possible intermediates along the non mevalonate pathway |
 |
Overall conclusions
The plant system M. citrata biosynthesises the
terpene linalyl acetate via the mevalonate independent pathway
Five deuterium atoms of DXP are retained during
plant terpene biosynthesis limiting the possible processes used to affect
de hydroxylation of DXP
Neither E.coli nor M. citrata possess enzymes
required to accept and phosphorylate neither known nor putative intermediates