Fluxmaximalisatie en convergentie in de evolutie
Wanneer we de diverse evolutiepatronen rondom ons bekijken, krijgen we soms de indruk dat er in de evolutie een tendens is naar "perfectie", of "voltooiing" in een welbepaalde richting, met als onderliggend doel steeds betere evolutieprocessen of organisatievormen af te leveren. In de oudheid werd deze perfectionerende drang veelal toegewezen aan de Almachtige God, die de touwtjes strikt in handen houdt, en door wikken en wegen de evolutie van het heelal in de juiste richting leidt. Vandaag de dag slaagt de mens erin om voor veel van deze schijnbare doelmatigheden, een plausibele verklaring te vinden, zonder dat hiervoor een oppermachtig wezen aan te pas komt.
Veelal zijn deze optimaliserende trends te verklaren door het feit dat er slechts een beperkt aantal oplossingen mogelijk zijn voor een bepaald probleem. Als gevolg hiervan, ontstaat een "convergentie" van oplossingen in de evolutie. Deze convergerende trends in de evolutie zijn ook te aanzien als "gewoontes" van de gemiddelde evolutie.
Daar waar de divergentie (beschreven in het vorige hoofdstuk) eerder een kenmerk was van de verspreiding in de evolutie, zullen we zien dat convergentie, en dus beperking van de verspreiding, eerder een kenmerk is van de fluxmaximalisatie (zie ook Figuur H9F1 voor de relatie tussen divergentie en convergentie).
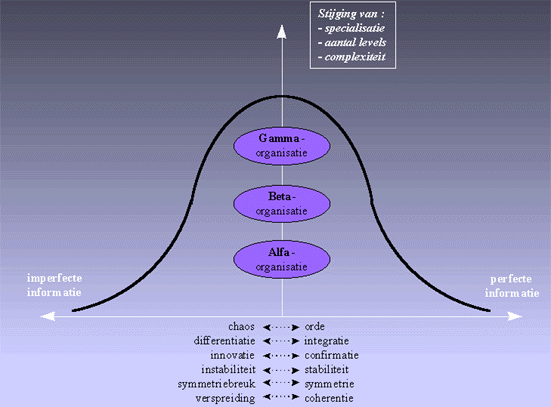
Figuur H10F1 : convergentie in de evolutie, balancerend tussen orde en chaos, innovatie en confirmatie, symmetriebreuk en symmetrie, instabiliteit en stabileit, afhankelijk van de "perfectie" van de informatie.
Deze trends zijn in sommige gevallen duidelijk waar te nemen, en in andere gevallen minder of niet.
We zullen aantonen dat drang om te evolueren naar meer complexe evolutiepatronen, afhankelijk is van de de kwaliteit van de informatie (de perfectie van de informatie). Zoals in bovenstaande grafiek is aangeduid, zal de kwaliteit van de informatie bepalen of er al dan niet een duidelijke richting in de evolutie is te onderscheiden. Hierbij zullen we zien dat de evolutie schippert tussen een schijnbaar streven naar perfectie, en een afglijden naar de totale verspreiding. Op dit continuum zullen we regelmatig een afwegen vinden van tegenstrijdige tendensen, zoals :- streven naar chaos of streven naar orde
- streven naar integratie of differentiatie
- innovatie of confirmatie
- instabiliteit of stabiliteit
- symmetriebreuk of symmetrie
- verspreiding versus coherentieWelke richting zal worden gevolgd in het netwerk van diverse mogelijke evoluties, is dikwijls afhankelijk van kleine en toevallige factoren. Opnieuw zullen we ons daarom beperken tot het aangeven van "gemiddelde" tendensen in de gemiddelde evolutie.
We zullen dit schipperen van de evolutie tussen enerzijds de perfecte informatie en anderzijds het volledig ontbreken van informatie, vanuit diverse standpunten bekijken. Elk van deze standpunten legt andere accenten, waardoor de lezer geleidelijk aan meer inzicht krijgt in wat men de "gestalt" van de gemiddelde evolutie kan noemen.
Terug naar de index van dit hoofdstuk
2. Innovatie versus confirmatie in de evolutie
Verspreiding is de basis voor alle variaties, en dus ook voor innovatie in de evolutie. Volledige verspreiding leidt echter niet tot nieuwe attractoren. Tijdens de evolutie ontstaat daarom een balans tussen innovatie en confirmatie.We zullen deze balans eens naderbij bekijken vanuit het standpunt van de creatie van "artificieel leven".
Bij het onderzoek naar de mogelijkheid om "artificieel leven" te creëren, is de vraag naar de essentie van een levend systeem van fundamenteel belang. Christopher Langton was één van de pioniers in dit onderzoek. Hij was er principieel van overtuigd dat levende systemen niks anders zijn dan fysische belichamingen van informatieverwerkende eenheden. Het verwerken van informatie is voor deze levende systemen nog belangrijker dan het verwerken van energie en materie. "Levende systemen gebruiken informatie om zichzelf op te bouwen, om voedsel te zoeken, om hun interne structuur te behouden,..." Langton concludeerde dat levende systemen gestuurd worden door manipulatie van informatie, en dat deze informatie-stroom de energie- en materiestromen op hun beurt bepaalt. In andere evolutiepatronen van het universum, is de rol van informatie minder dominant, en overheerst de tendens naar verspreiding. Deze tegenstelling intrigeerde Langton in die mate, dat hij een virtueel experiment ontworp om het gedrag van systemen onder invloed van diverse informatiestromen te onderzoeken. Hij ontwierp in zijn computerwereld een aantal programma’s voor artificieel leven, die enkel verschilden in de mate waarin informatie vrij kan vloeien doorheen de informatietabellen die zijn "cellulaire automata" aanstuurden. De mate van vrije vloei van informatie kon hij aanpassen met een parameter die hij de "lambda-parameter" doopte.
Het programma was zo opgebouwd, dat hij zijn lambda parameter kon laten variëren tussen nul en één.
Wanneer hij de programma’s liet draaien op de computer, deed hij een aantal opmerkelijke vaststellingen.
Als de lambda-waarde dicht bij nul was, was de vrije vloei van informatie heel laag. De informatie is als het ware bevrozen : ze blijft lang geconserveerd, doch er is weinig verandering waar te nemen. Net als bijvoorbeeld de watermoleculen in ijs nauwelijks bewegen (met uitzondering van de locale trillingen), en er in het ijs weinig veranderingen plaatsgrijpen. De onderlinge verhoudingen, en dus de orde tussen de watermolecules, blijft onveranderd. De informatie verandert nauwelijks, of met andere woorden, er is weinig vernieuwing of innovatie waar te nemen in het systeem : de actuele toestand van het systeem blijft geconfirmeerd.
Wanneer daarentegen de lamda-waarde zeer hoog is (dicht bij 1), kan de informatie zeer vrij vloeien. Er heerst chaos binnen het systeem, en het aantal veranderingen is ontelbaar. De inwendige orde in het systeem wordt voortdurend verstoord. Het systeem is zodanig innoverend, dat er geen of nauwelijks geen stabiele evolutiepatronen kunnen waargenomen worden. In fysische termen is deze toestand te vergelijken met een gas, waarbij de molecules chaotisch bewegen, en waar enkel een gemiddelde symmetrie is waar te nemen, doch geen stabiele evolutiepatronen met een duidelijk waarneembare individuele symmetrie.In geen van beide condities, kon Langton evolutiepatronen waarnemen, die de zelforganiserende kenmerken van artificieel leven in zich dragen.
Tussen beide uitersten bestond echter een gebied, waar de stroming van informatie snel was, doch niet zo snel dat de relatie met de toestand in elk van de voorgaande toestanden volledig verbroken is. Dit gebied heeft zijn fysisch equivalent in een vloeistof. Het is precies in dit gebied van een beperkte vrije vloei van informatie, dat artificieel leven kan aangetoond worden. Artificieel leven van het type dat reeds eerder werd beschreven in het spel "Leven", heeft een lambda-waarde van ongeveer 0.27. Er kon aangetoond worden dat complex zelforganiserende gedrag, enkel voorkomt in bepaalde gebieden van de lambda-waarde.
Langton kon dus met zijn experiment aantonen dat het complexe zelforganiserende gedrag (waarvan het leven een bijzondere vorm is), zich enkel kan voordoen in een gebied waar informatie enigzins vrij kan stromen, doch ook niet zo vrij dat het continu met het verleden breekt. Een hoge complexiteit doet zich enkel voor op een grensgebied tussen orde en chaos, waar een balans gevonden wordt tussen innovatie en confirmatie van de informatiestroom. Dit is symbolisch voorgesteld in de klokvormige curve die we in figuur 1 hebben weergegeven.
Bij levende systemen zoals bijvoorbeeld een boom, een koe of een maatschappij, bemerken we inderdaad subsystemen die gekenmerkt zijn door een relatief snelle doorstroming van materie en/of informatie (bijvoorbeeld de kanalen voor de sapstroom in een boom, het zenuwstelsel van de koe, of een communicatienetwerk van een willekeurige maatschappij), terwijl andere een eerder stabiel karakter vertonen. Zo zullen de ondersteunende structuren van deze levende systemen (de beenderen bij de koe, de stam van de boom, en bijvoorbeeld de wegeninfrastructuur in een maatschappij) relatief traag veranderen.
De balans tussen innovatie en confirmatie is van cruciaal belang voor het opbouwen van dynamische systemen met hoge complexiteit.Terug naar de index van dit hoofdstuk
Systemen met een hoge complexiteit, zoals Gamma-interactoren, zijn gekenmerkt door een grote hoeveelheid informatie nodig voor de beschrijving van het ontwikkelingsproces. Tijdens de ontwikkeling van een Gamma-interactor, is de kans op het optreden van foutieve informatie (mutaties in de informatie) reëel. Ook bij de replicatie van de informatie in de Gamma-attractor kunnen fouten optreden, die uiteindelijk zullen leiden tot afwijkingen in vergelijking met het normale evolutiepatroon. Globaal gezien zijn er 2 mogelijke manieren waarbij de kans op het optreden van fouten kunnen verhinderd of beperkt worden. Een eerste methode is het gebruik van redundante informatie en een tweede is het gebruik van controlesystemen.
- Redundante informatie
Informatie vertoont redundantie als hun beschrijving of codering niet de kortst mogelijke is, met andere woorden, als er andere beschrijvingen of coderingen mogelijk zijn die met een kleiner aantal bits dezelfde betekenis weergeven.
Redundantie informatie komt in ons dagelijks leven veel voor. De menselijke hersenen zijn immers vergeetachtig, en informatie gaat verloren in ons geheugen. Dit is nog meest het geval bij kinderen, die vele dagelijkse activiteiten nog niet als een routineproces in hun geheugen hebben opgeslagen. Om die reden moet talrijke redundante informatie gegeven worden zoals "vergeet je tanden niet te poetsen ; goed opletten als je de straat oversteekt, en dergelijke ...". Electronische geheugens vertonen dit euvel veel minder, en de hoeveelheid redundante informatie die moet opgeslagen worden is dan ook veel kleiner.
Elke taal bevat in min of meerdere mate redundantie. Het Nederlandse woord "geeuwen" bevat een aantal overtollige letters ; het woord "gewen" zou immers even goed dezelfde informatie weergeven. De letters "e" en "u" in het midden zijn dus redundant. De verschillende menselijke talen zijn door taaldeskundigen bestudeerd naar het voorkomen van redundantie. Zo bevatten Duitse woorden gemiddeld 66% redundante letters, en de Engelse taal 75%. Dit betekent dat een groot deel van de letters in het Duits en het Engels, niet wezenlijk bijdragen tot de informatie-overdracht. Het Engelse woord "through" zou bijvoorbeeld kunnen herleid worden tot "tru", waarbij heel wat redundante letters vermeden zijn. Het nadeel van het weglaten van alle redundante letters in een taal, zou kunnen zijn dat bij het copiëren of lezen van teksten, meer fouten gemaakt kunnen worden, waardoor de teksten moeilijker leesbaar worden.
Geschreven teksten worden dikwijls aangevuld met grafieken, foto’s en referenties, die de leesbaarheid en begrijpelijkheid van het geheel verhogen, doch in essentie veelal redundant zijn.
Niet enkel bij geschreven teksten, doch ook bij bijvoorbeeld gesproken presentaties wordt veelvuldig gebruik gemaakt van redundante informatie. De spreker gebruikt naast zijn verbale taal, ook een niet-verbale lichaamstaal om zijn presentatie kracht en emotie bij te brengen. Bovendien wordt gebruik gemaakt van bijvoorbeeld dia’s of een video om de informatieoverdracht maximaal te ondersteunen. Op regelmatige tijdstippen wordt ook een herhaling gegeven van de belangrijkste boodschappen, om het voorkomen van fouten bij de informatie-overdracht te beperken. De gesproken taal op zich bevat vele redundanties. De meeste klanken duren immers langer, dan echt nodig is om herkenbaar te zijn via het menselijke oor. Samen met al dan niet bewuste pauzes in het spraakpatroon, maken ze het de toehoorder makkelijker om de presentatie in al zijn details te begrijpen.
De menselijke communicatie is dus een voorbeeld bij uitstek van het gebruik van redundante informatie.
De mate waarin de informatie als redundant wordt beschouwd, is natuurlijk ook afhankelijk van de ontvanger (bijvoorbeeld de toehoorder) van de informatie, die de interpretatie doet. Voor een baby is nagenoeg alle informatie nieuw, en bevat de boodschap geen enkele redundantie. Voor een "alwetende", is elke informatie bekend, en dus redundant. Vandaar dat ook het begrip "leren" soms vertaald wordt als "het ontdekken en benutten van redundanties". Wanneer redundanties ontdekt worden, kan men tot een kortere, meer cryptische omschrijving van de interactor komen. Dit kan bijvoorbeeld door de regelmaat in het patroon te ontdoen van zijn herhalingen, door de symmetrie in een wiskundige formule te gieten. Zo geeft een wiskundige fractaal een zeer goede, doch cryptische beschrijving van soms prachtige interactoren in de natuur. De wiskundige formulatie is echter enkel begrijpelijk voor de specialist, die vertrouwd is met de redundantie van de beschrijving. Van zodra de redundanties "ontdekt" zijn, kunnen ze verder benut worden voor het bouwen van modellen of onderzoeken van interactoren. Tot voor het periodiek systeem der elementen in de chemie ontdekt was door Mendeljev, was de ontdekking van elk nieuw atoom een volledige stap in het onbekende. Mendeljev heeft de wetmatigheden en regels van de bouw en structuur van atomen ontdekt, en in een periodiek systeem weergegeven dat de redundantie beperkt. Op die manier kan het bestaan van nieuwe, nog niet ontdekte atomen niet enkel voorspeld worden, doch ook hun eigenschappen kunnen voorspeld worden. Elk theoretisch model dat wordt opgebouwd, start vanuit de redundante regelmaat die ontdekt is. De volgende stap is het beperken van de redundanties, door het geven van een meer cryptische beschrijving van de werkelijkheid.
Zeer regelmatige systemen bevatten zeer veel redundanties. In het kristalrooster worden de onderlinge posities immers telkens opnieuw herhaald. Deze redundantie van positionele informatie, met andere woorden de regelmaat in het kristal is ook de basis van de macroscopische symmetrie. Symmetrie wijst altijd op een zekere mate van redundantie : zonder herhaling is er immers geen symmetrie. De tweezijdige symmetrie van de mens en vele andere zoogdieren, is te wijten aan de aanwezigheid van één redundant oor, oog, neusgat, één redundante arm en been. De afwezigheid van deze ledematen of zintuigen maakt het leven immers een heel stuk moeilijker, en als er van elk twee zijn, is het verlies van één ervan minder kritisch. Deze organen en ledematen konden wellicht ook op niet symmetrische wijze over het lichaam verspreid worden. Dit zou echter evolutionair gezien minder efficiënt zijn, zowel tijdens de ontwikkeling als tijdens het routinegebruik. (Stappen met één been op het hoofd en een been op het bekken is wellicht niet zo evident). Spontane symmetrie is dus een uiting van van efficiënte en effectieve redundantie. Interessant hierbij is op te merken dat de geslachtsorganen niet in tweevoud bestaan ; ze zijn niet zo kritisch voor het overleven van het individu, doch des te meer voor het overleven van de soort. Belangrijker nog dan het geslachtsorgaan is de ontwikkelingsinformatie. De redundantie hiervan is dan ook zeer groot. Elke man produceert enorme hoeveelheden zaadcellen, die formele (over)dragers zijn van de ontwikkelingsinformatie. Bovendien is de informatie in het DNA dubbel aanwezig : de twee strengen van de "dubbelhelix" bevatten immers dezelfde informatie. Een grote hoeveelheid redundante informatie is een kenmerk dat we moeten situeren aan de rechterhelft van grafiek 1. Een kleine hoeveelheid redundantie, waarbij elke situatie nieuw en moeilijk voorspelbaar is, ligt duidelijk aan de andere kant van de grafiek. Naarmate systemen echter complexer worden, zullen meer en meer redundanties nodig zijn om fouten te beperken. Zoals tijdens het ontwikkelen van een baby meer en meer redundanties bekend en benut worden, zo worden in de evolutie van andere complexe evolutiepatronen "redundante bouwstenen" gebruikt als stabiele basis voor nieuwe informatie. De evolutiepatronen hebben baat bij aan de ene kant een voldoende hoeveelheid redundantie, om de stabiliteit van het systeem te waarborgen (onder andere door minder fouten), doch anderzijds ook voldoende nieuwheid van informatie, om de groei en het aanpassingsvermogen te garanderen. Organisaties die volledig vastgelegd zijn in redundante informatiestromen (procedures, wetgeving), vertonen immers het gevaar niet flexibel genoeg te kunnen inspelen op snelle veranderingen in de omgeving. Dit is de reden waarom zogenaamde "break-through" innovaties vaak ontwikkeld zijn door kleine, flexibele bedrijfjes, terwijl grote en logge organisaties soms zover doorgaan op successen uit het verleden, dat ze met hun grote inertie afstevenen op een dalende curve in hun levenscyclus (in het Engels zegt men "core comptencies can become core rigidities"). Op dezelfde wijze hebben bijvoorbeeld bacteriën en virussen een veel groter aanpassingsvermogen dan bijvoorbeeld dinosaurussen.
Redundante informatie waarborgt dus het behoud van de regelmaat (de confirmatie) in complexe organisaties, terwijl het ontbreken ervan precies de vernieuwing en aanpassing (de innovatie) van organisaties en evolutiepatronen ten goede komt. Complexe systemen hebben beide elementen nodig om hun overleving op lange termijn te waarborgen.- Controlesystemen en vorming van cycli
Redundantie is een hulpmiddel om fouten bij informatieoverdracht te beperken en de correctheid van de informatiestroom in complexe systemen te waarborgen.
Naarmate de complexiteit van systemen echter hoger wordt, zal het aantal variaties in de omgeving zo groot worden, dat het ontstaan van controlesystemen noodzakelijk wordt. Deze zorgen ervoor dat verstoringen in de omgeving beperkt worden, en dat het evolutiepatroon zijn "steady-state"-stromen van energie, materie en informatie kan behouden.
Om deze verstoringen te kunnen compenseren, moet het controlesysteem weten welke mogelijke acties er zijn (of voorkennis hebben) over de mogelijk compenserende acties op de verstoring. Dit impliceert dat het een model over de omgeving of zichzelf moet bevatten. Ashby stelde in 1970 "A good regulator of a system, must be a model of that system". Om een model op te bouwen over een systeem (bijvoorbeeld de omgeving), moet dit systeem ontdaan worden van zijn redundanties, zodat een cryptische beschrijving kan worden bekomen (zoals een attractor cryptische informatie bevat met betrekking tot de interactor). De mate van crypticiteit kan hierbij sterk verschillen. Als we bijvoorbeeld bepaalde bacteriën plaatsen in een voedselarme omgeving, zullen ze met hun trilhaartjes rond hun lichaam beginnen trillen. Hierdoor zullen ze zich op blinde wijze verplaatsen, waarbij de verplaatsing hopelijk (maar niet noodzakelijk) naar een meer voedselrijke omgeving leidt. Het model in deze bacteriën is van het type :
- "als er weinig voedsel is, laat trilhaartjes bewegen
- als er veel voedsel is, laat trilhaartjes rusten".
Dit model is eenvoudig en weinig cryptisch, doch zal ook niet altijd tot het gewenste resultaat leiden.
De strategie die een vleermuis gebruikt om zijn prooi te vangen is al een stuk gesofisticeerder. De vleermuis zendt een ultrasoon geluid uit, met een frequentie van 40000 tot 200000 Hertz, die ver boven de waarneembare frequentie voor de mens ligt (40 tot 20000 Hertz). Hiermee localiseert hij obstakels (en bepaalt zo zijn vliegweg) en mogelijke prooien op basis van het type weerkaatsing van deze geluidsgolven. Het heeft lang geduurd vooraleer de mens deze strategie heeft ontdekt. Eénmaal echter dat dit model bekend is geraakt, heeft de mens het toegepast om bijvoorbeeld in boten te gebruiken (in de sonar’s) voor het localiseren van obstakels of vijandige bewegingen onder water.
Complexe interactoren bevatten tal van zelfregulerende modellen. Deze regulaties zijn typische kenmerken van zelforganiserende Beta-systemen.
In veel gevallen gaat het om homeostatische regulaties, die de verstoringen van buitenaf of de stresscondities trachten te minimaliseren. Hierbij wordt "Het principe van Le Chatelier" gebruikt : dit principe is gegroeid vanuit de chemie, doch luidt in algemene termen als volgt :
"Elk stabiel systeem dat onder stresscondities wordt geplaatst, zal in een richting evolueren die de stress tracht te minimalizeren." Als een dier honger heeft, zal het eten zoeken ; als een dier te koud heeft, zal het zich verplaatsen naar een warmere plaats, of beginnen rillen (om zelf meer warmte te produceren). Als een mens teveel informatie ("informatie overload") krijgt (bijvoorbeeld een manager in een onderneming) zal hij samenvattingen van de informatie vragen, of meer selectief of sneller de aangeleverde informatie doornemen. Krijgt hij te weinig informatie, zal hij bij zijn ondergeschikten de informatie opvragen, of desnoods gebruik maken van externe experten of consultants voor het verzamelen van de informatie. Het principe is dus geldig voor excessen of tekorten in de stromen van informatie, materie en energie. Elk regulerend model tracht ervoor te zorgen dat de M/E/I-stromen binnen het bereik blijven waarvoor het ontworpen is. Lukt dit niet, dan verliest het subsysteem de controle. In dit geval kan de controle overgenomen worden door subsystemen die hoger in de hiërarchie staan, of die ontworpen zijn voor deze noodsituaties (een thermostaat regelt de temperatuur in de huiskamer, doch bij brand roepen we de brandweer). In veel gevallen kan dit echter de ondergang van de systemen betekenen. Deze werkzame gebieden van de controlesystemen zijn soms verbazend klein. De meeste fysiologische controlesystemen van de mens werken best bij 37°C ; bij een temperatuur die enkele graden hoger ligt is de mens ziek, en een lichaamstemperatuur van tien graden hoger betekent de dood.
In veel gevallen wordt hierbij gebruik gemaakt van feedback-cycli en hypercycli.
Het principe van de fluxmaximalisatie is op zich ook een regulerend systeem : het beperkt immers de kans dat gradiënten van M/E/I worden opgebouwd. Hoe groter de gradiënten zijn, hoe moeilijker het wordt om de gradiënt nog verder te verhogen.
Hoeveel modellen een complex systeem ook bevat, en welk regulerend principe ook gebruikt wordt, de modellen zullen nooit perfect zijn. Elk model is slechts een cryptische afspiegeling van de werkelijkheid, die de volledig diepte van de realiteit nooit kan evenaren. Dat deze stelling klopt, kan wellicht niet bewezen worden, doch kan wel als volgt worden ingezien. Stel dat het model beter is dan de werkelijkheid die het weergeeft, dan zou de simulatie in het model meer tijd nodig hebben om te reageren dan het echte proces, waardoor anticipatie onmogelijk wordt.
Deze traagheid van de modellen kan bijvoorbeeld waargenomen worden als de modellen gebaseerd zijn op feedback systemen. Deze hebben de neiging te oscilleren naar hun evenwichtswaarde. Wanneer bijvoorbeeld een organisme uit evenwicht geraakt, zal het rond zijn evenwichtpositie schommelen. De reden is dat de informatieoverdracht in het zenuwstelsel slechts reageert met een snelheid van enkele honderdsten van een seconde, wat te traag is om perfect evenwicht te garanderen. Het model is dus niet perfect. Analoog kan het dagen, weken of maanden duren, vooraleer een organisatie een fout heeft hersteld, bijvoorbeeld een herstelling in een assemblagelijn voor auto’s, het terugroepen van een medicijn van de markt omdat het onveilig is of het corrigeren van een managementstrategie. Hoe complexer het systeem, hoe trager de feedback. Een maatschappij bijvoorbeeld reageert in regel heel traag op signalen die gegeven zijn tijdens de voorgaande verkiezingen.
Hoe correcter het model, hoe trager de bijsturing. Elk model moet dus een balans vinden tussen juistheid van informatie en snelheid van informatiestroom. Doordat het model imperfect is, is er ruimte voor variaties, en is er ruimte voor innovatieve veranderingen. Opnieuw vinden we dit evolutionair paradigma terug. Elk model in complexe interactoren is imperfect, wat op korte termijn tot de ondergang van de interactor kan leiden ; doch op lange termijn is deze onvermijdelijke imperfectie ook de beste garantie voor het voortbestaan van het complexe systemen, omdat het de enige mogelijkheid is om voor onvoorzienbare stresscondities toch een geschikt gedrag te ontwikkelen. Complexe systemen dansen door de beperktheid van hun regulerende modellen op een dunne koord tussen innovatie en confirmatie, doch deze koord is ook de enige weg naar overleving in de verre toekomst.
Door controlesystemen kan de blinde verspreiding dus nooit volledig worden beperkt ; doch dit is juist de beste garantie voor het ontwikkelen van verbeteringen aan de modellen en de evolutiepatronen.Terug naar de index van dit hoofdstuk
2.3. Innovatie en confirmatie in een ecosysteem
De meest complexe systemen op onze aardbol zijn de ecosystemen. Deze ecosystemen zijn het resultaat van de interactie tussen biologische, fysische en chemische componenten in de natuur, die samen geïntegreerd functioneren. Alfa- Beta, en Gamma-interactoren ageren samen in harmonie in een ecosysteem.
Daarom is het interessant om na te gaan hoe de balans tussen innovatie en confirmatie in een ecosysteem wordt gehanteerd. Om dit te doen, heeft Eric Schneider een vergelijking gemaakt tussen een jong ecosysteem, en een matuur ecosysteem. De onderstaande tabel is gebaseerd op enkele van de conclusies.
Tabel H10T1 : Vergelijking tussen een jong en een matuur ecosysteem.
Ecosysteem kenmerk
Jong ecosysteem
Matuur ecosysteem
Structuur
- kleine interactoren (bijvoorbeeld planten en bomen)
- eenvoudige structuren
- weinig hiërarchische niveau’s
- kleine diversiteit
grote interactoren complexe structuren veel hiërarchische niveau’s grote diversiteit Process
- minder goede captatie van de instroom van M/E
- hoge specifieke entropieproductie
- lage totale entropieproductie
- lage totale M/E/I flux
- eenvoudige cycli
- hoge systeemefficiëntie
- snelle reactietijd
goede captatie van de instroom van M/E lage specifieke entropieproductie hoge totale entropieproductie hoge totale M/E/I-flux (uiting van fluxmaximalisatie) veel en complexe cycli, veelal met recyclage (weinig verspilling) lage systeemefficiëntie trage reactietijd Informatie
- relatief meer verspreiding van de M/E stroom
hogere coherentie van de M/E stroom Selectiekenmerken
- "r"-selectie (snelle groei)
- brede ecologische niches
"K"-selectie (trage groei) smalle ecologische niches (doorgedreven specialisatie)
Het is duidelijk dat in een jong ecosysteem de "innovatie" de bovenhand heeft, terwijl in een matuur ecosysteem de "confirmatie" de bovenhand heeft. Het is ook duidelijk dat de evolutie van een jong naar een oud ecosysteem, een mooie illustratie is van vele kenmerken van "de gemiddelde evolutie".
Eén van de eerste diepgaande studies van deze ecologische successie in natuurlijke systemen is gedaan door Henry Thoreau in een lezing genaamd "The succession of forrest trees". Deze lezing dateert van 1860. Hij stelde vast dat de natuur een geordend proces van plantenontwikkeling vertoont, waarbij er een voorspelbare sequentie optreedt van de ene soort planten door de andere. Als bijvoorbeeld een grasland vrij wordt gelaten, zal het eerst langzaam evolueren naar een dennebos, om finaal te eindigen in een loofbos met eiken. Dit is een proces dat ongeveer 150 jaar duurt. In dit proces van evolutionaire successie, worden bestaande dominante attractoren vervangen door verbeterde dominante attractoren.
Deze vaststellingen van Thoreau dateren ongeveer van dezelfde periode als de publicatie van Darwin’s "Origin of species". Darwin stelde hierbij vast dat natuurlijke selectie het resultaat is van organismen die met elkaar concurreren voor beperkte grondstoffen.
Later werd hierbij wetenschappers toegevoegd dat het effect van deze constante competitie is dat planten en dieren evolueren naar een grotere efficiëntie.
In 1968 vervolledigde Margalef deze vaststellingen met diepgaandere wetenschappelijke gegevens.
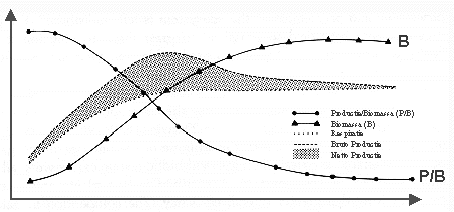
Deze evolutie is verduidelijkt in onderstaande figuur.
Figuur H10F2 : biologische productie-indicatoren gedurende de groei en ontwikkeling van een ecosysteem (naar E. Schneider, 1988)
Margalef concludeerde dat in een biologisch ecosysteem de ecologische successie (de evolutie van een jong naar een matuur ecosysteem) een zelf-organiserend systeem is, dat evoleert naar een minimale entropieproductie per éénheid. Margalef stelde voor de energieflow per eenheid biomassa (de "P/B-ratio" : dit is maat voor de entropieproductie per eenheid biomassa) te gebruiken als maat voor de efficiëntie van het ecosysteem. Uit metingen blijkt dat naarmate het ecosysteem matuurder wordt, de P/B ratio daalt (zie figuur) wat de hypothese bevestigt. De totale biomassa (B) van het ecosysteem stijgt gedurende de ontwikkeling van het ecosysteem.
Beide vaststellingen komen overeen met het principe van de fluxmaximalisatie.
Bij de evolutionaire successie zullen nieuwe biologische individuen bestaande vervangen, als ze per eenheid energie efficiënter zijn, en dus relatief minder entropie produceren. Op deze wijze wordt de verspreiding van energie per individu meer vertraagd, en kan meer energie worden opgeslagen onder de vorm van informatie of structuur, wat de complexiteit doet toenemen. In totaal zal echter de entropie-productie stijgen, omdat de totale biomassa relatief sterk toeneemt. De totale respiratie – als maat voor de entropie-productie - blijft ook stijgen gedurende de evolutie van het systeem (cfr. figuur 2)
Mature ecosystemen hebben relatief lange levenscycli, en veelal vrijwel gesloten materiaalcycli, in tegenstelling tot jonge ecosystemen. Bij een jong ecosysteem is de biomassa immers laag in verhouding tot de draagkracht van de omgeving, waardoor er nog weinig competitie is voor de grondstoffen. Pas later in de evolutie van het systeem worden de grondstoffen een beperkende factor, en hebben meer efficiënte soorten een competitief voordeel.
Dit betekent dat de maximale flux van het ecosysteem, aanvankelijk weinig beperkt is door de draagkracht van de omgeving, waar dit in een latere fase wel het geval is. De specifieke entropieproductie is daardoor bij een jong ecosysteem hoog, terwijl dit lager is bij een meer matuur ecosysteem. De totale entropieproductie echter, is hoger bij het het mature ecosysteem, precies door de hogere efficiëntie. De hogere diversiteit aan soorten in een matuur systeem, is hierbij niet alleen verantwoordelijk voor deze hoge entropieproductie (alle mogelijke voedselniches worden benut), doch ook voor de hogere complexiteit.
Binnen het ecosysteem zijn de verschillen tussen de soorten op hogere en lagere hiërarchische niveau’s treffend. De soorten die hoger staan in de hiërarchie, kunnen in het algemeen een grotere "bandbreedte" van voedsel en informatie benutten, terwijl de lagere soorten gedwongen worden zich meer te specialiseren in beperkte voedsel- en informatie-niches. De interactoren aan de top van de piramide zijn eerder "generalisten", terwijl deze aan de basis eerder "specialisten" zijn. Hoe matuurder het ecosysteem, hoe meer de beschikbare energie-gradiënt (het voedsel) gedempt wordt, door de betere benutting. De soorten hoger in de hiërarchie zijn betere gradiëntdempers (K-selectie) dan de soorten lager in de hiërarchie (r-selectie). Ze kunnen over het algemeen echter trager inspelen op gradiëntwijzigingen dan soorten lager in de hiërarchie. Een aantal kenmerken van meer mature ecosystemen vinden we dus eerder terug bij soorten die hoger in de hiërarchie staan, terwijl deze van jonge ecosystemen eerder terug te vinden zijn bij de soorten die lager in de hiërarchie terug te vinden zijn.
Lionel Johnson vergelijkt het biosysteem met een enorme informatieverwerkende processor. Hij verwoordt dit heel krachtig als volgt :"Het ganse biosysteem fungeert als een informatie extractie- en verwerkingseenheid, in dewelke de verschillende informatieverwerkende eenheden functioneren als een serie van gekoppelde schokdempers, die een toenemende demping tot gevolg hebben, op elk bijkomend niveau in de hiërarchie."
Kay en Schneider hebben de verschillen tussen ecosystemen nog op een indirecte wijze bevestigd door vergelijking van het regenwoud in de Amazone, gematigde vlakten in centrum en het Oosten van de Verenigde Staten en de Sahara-woestijn. Deze drie gebieden hebben volkomen verschillende ecosystemen, aangepast aan het klimaat. Door metingen in de lucht in verband met de warmtetransfer in deze systemen, kon vastgesteld worden dat in de Sahara, de invallende zonne-energie grotendeels wordt gereflecteerd in de lucht, wat betekent dat het ecosysteem in zijn geheel nauwelijks een metabolisme heeft opgebouwd, om de invallende zonne-energie te capteren en te verwerken. In een regenwoud daarentegen is een zeer rijk en matuur ecosysteem opgebouwd, dat heel veel van de binnengevallen zonne-energie degradeert tot een lagere energievorm. De M/E stroomt wordt op meer coherente wijze ge-organiseerd, wat wijst op een hogere complexiteit en meer informatie om de M/E stromen in coherente banen te leiden. Een tropisch regenwoud is dus een efficiënter systeem, met een hogere totale energieflux en een hogere gradiënt-dissipatie, door het geheel van kenmerken dat opgesomd is in de rechterkolom van tabel 1.In elk geval kunnen we uit deze vergelijking van ecosystemen leren dat gedurende de ontwikkeling en veroudering van het ecosysteem, de confirmatie belangrijker wordt en de innovatie daalt. Hierbij stijgt de totale energieflux in het systeem (fluxmaximalisatie), daalt de verspreiding per eenheid biomassa en stijgt de totale verspreiding. De hogere energieflux komt overeen met een betere interne organisatie. In elk van de ecosystemen dient echter een balans te worden gerespecteerd, die overeenkomt met de beschikbare energie-gradiënt en de draagkracht van de omgeving.
Deze kenmerken zijn min of meer typerend voor de ganse evolutie "van bit tot atoom en ecosysteem".Terug naar de index van dit hoofdstuk
3. Symmetrie en symmetriebreking in de evolutie
Symmetrie is een eigenschap die natuurkundigen nu méér bezighoudt dan ooit.
Het onderzoek naar de meest elementaire eenheden in ons universum is een onderzoek naar "supersymmetrieën", een essentieel kenmerk van het prille universum. Hiervan zijn de supersymmetrische "super"snaren een belangrijk onderdeel. De basisidee is dat de hele fysische natuur zou beheerst worden door één "super"kracht, die de eigenschappen in zich draagt om het heelal te doen ontstaan, en de oorzaak te vormen voor het bestaan van materie en energie, en alle afgeleide "universele" Alfa-attractoren. Voor het onderzoek naar passende wiskundige beschrijvingen, is de toetsing van de symmetrische kenmerken een belangrijk criterium voor de kwaliteit van de gevonden formules.Deze belangstelling voor symmetrieën heeft in de loop van de geschiedenis ups en downs gekend.
Al sinds de oudheid is de mens geboeid geweest door de natuur rondom zich heen. Sinds het onstaan van het menselijk bewustzijn zijn de optimale vormen van planten en dieren, en de indrukwekkende harmonieën van de hemellichamen als een teken aanzien geweest van de "goddelijke aanwezigheid"."Wat anders dan een goddelijke aanwezigheid kan immers de talrijke "optimale ontwerpen" rondomons verklaren ? " is de indirecte vraag die vele generaties zich stelden, en waar de mens een steeds beter antwoord op tracht te formuleren. Deze zoektocht naar een verklaring voor deze optimalisatie en symmetrievorming in de evolutie, gaat historisch gepaard met de zoektocht naar de wetmatigheden die hiervoor aan de basis liggen.
In de volgende paragrafen zullen we aantonen dat in de gemiddelde evolutie een evenwicht gevormd wordt tussen symmetrievorming (als kenmerk van "optimale vormen") en symmetriebreuk.
Aangezien informatie aan de basis ligt van coherentie, en coherentie een basis is voor symmetrie, zullen we een aantal voorbeelden geven van symmetrie en symmetriebreuk voor Alfa-, Beta- en Gamma-attractoren.Terug naar de index van dit hoofdstuk
Ga naar het vervolg op de volgende pagina