3.1. Symmetrie en Alfa-informatie
- Een historische terugblik
De oude grieken vertoonden een groot ontzag voor optimale vormen zoals cirkels en bollen. Hun "volmaakte symmetrie" was een uiting van het goddelijke. Om die reden geloofden ze dat planeten zich rond de aarde voortbewegen in een cirkelvormige baan (de optimale beweging) . De filosoof Xenophanes ging zelfs zover, dat hij God de vorm van een bol toekende. God moest immers vanuit alle standpunten gelijk zijn en volmaakt homogeen, en dit kon enkel indien Hij de vorm van een bol zou hebben. Aanvankelijk geloofden ze dat de aarde een platte schijf was, die omringd werd door oceanen en overspannen door een bolvormige koepel. Pythagoras was echter reeds overtuigd dat de aarde bolvormig was, en Erastosthenes berekende zelfs dat de omtrek van de aarde rond de 37000 km moest liggen. Dit is kennis die tijdens het christendom in Europa in het volgende millenium weer verloren was gegaan : uit de bijbel kon immers afgeleid worden dat de aarde plat was....
De voorliefde van de oude Grieken voor de bolvorm en cirkelvormige patronen als basisvormen voor het heelal, werden echter weer actueel bij de ontdekkingsreizen in de 15de eeuw, toen de Renaissance de oude Griekse geschriften in ere herstelde. Voor deze ontdekkingsreizen was aardrijkskunde, en het vinden van de korste of vlugste oversteek van cruciaal belang voor het welslagen van deze avontuurlijke missies.
In de volgende eeuwen werd verder gezocht naar verklaringen voor de optimale en symmetrische vormen in de natuur. Eén van de eerste belangrijke pogingen voor een algemeen verklarend grondbeginsel werd gedaan door de Pierre-Louis de Maupertuis (1698-1759). Zijn algemeen "metafysisch principe" luidde als volgt :
Als er in de natuur een verandering plaatsgrijpt, dan is de hoeveelheid actie die nodig is voor deze verandering, zo klein mogelijk.
Maupertuis was overtuigd dat zijn algemeen principe, dat zowel voor in rust verkerende als bewegende objecten geldig was, een overtuigend bewijs was van het bestaan van God. De algemene formulering van dit "principe van de kleinste werking", werd later door Gottfried Wilhelm Leibniz verder verduidelijkt in zijn belangrijktste werk : "Théodicée : sur la bonté de Dieu, la liberté de lhomme et lorigine du mal", dat in 1710 werd gepubliceerd. In dit werk verdedigde onder andere de twee volgende stellingen : ten eerste dat ons heelal het beste is van alle mogelijke ontwerpen, en ten tweede dat indien er toch suboptimale kenmerken zijn waar te nemen, dat deze toch deel uitmaken van de door God vooraf gewilde harmonie. Aangezien God immers Almachtig en Alwetend is, zou Hij, indien dit ook maar mogelijk is, een perfecte wereld scheppen. Als er dan toch onvolmaakte kenmerken zijn aan onze wereld (zoals kwaad onder de mensen, natuurrampen,...) dan is dit omdat het onmogelijk was voor God om een nog betere wereld te scheppen.
Hoewel Leibniz veel aanhangers had, waren er ook in zijn tijd veel tegenstanders. Zijn bekendste was wellicht Voltaire, die als antwoord op het werk van Leibniz een scherpe satire schreef, met als titel "Candide, ou lOptimisme.
Op het einde van de zeventiende eeuw kwam echter de doorbraak in de wiskunde, die het zoeken naar optimale vormen, wegen, ... met een mathematische precisie zou toelaten. Onafhankelijk van elkaar ontdekten Newton en Leibniz de infinitesimaalrekening, die de basis zou vormen voor de variatierekening.
- Symmetrie van (Universele) Alfa-organisaties
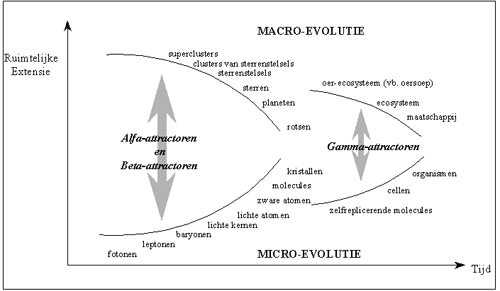
Nemen we bijvoorbeeld de gravitatiekracht als universele Alfa-attractor. Elk lichaam trekt een ander lichaam aan, met een kracht die evenredig is met het product van zijn eigen massa, vermenigvuldigd met de massa van het andere lichaam (cfr. de universele gravitatiewet van Newton).
Doordat elk deeltje van het lichaam een evenredige bijdrage heeft tot de gravitatie, ligt het zwaartepunt (bij een homogeen lichaam) in het geometrisch centrum van het lichaam. Alle lichamen die rond dit lichaam aanwezig zijn, worden als het ware aangetrokken tot het zwaartepunt. Om die reden is de zwaartekracht in elk lichaam tot een "puntattractor" te herleiden. Trekt een lichaam heel veel kleine deeltjes rondom zich aan, dan zullen deze zich homogeen rond de puntattractor verspreiden, waardoor we deze interactor zullen waarnemen als een bol.
Het feit dat de gravitatiekracht de kenmerken vertoont van een puntattractor, is de oorzaak van de vele bolvormige symmetrieën die we waarnemen in Alfa-interactoren.
Door omgekeerd te redeneren, kunnen we stellen dat de vorm van de interactor, iets zegt over het type attractor dat het heeft tot stand gebracht. Het feit dat de aarde (bij eerste benadering) een bolvorm vertoont, zou ons op basis van deze redenering al doen vermoeden dat een puntattractor aan het werk is.
Uit onze natuurkunde herinneren we dat we alle "universele krachten" of "universele attractoren" kunnen voorstellen als punten, wat inderdaad aangeeft dat het allen punt-attractoren zijn, die normaliter bolvormige evolutiepatronen impliceren. Zo worden de neutronen en de protonen in de atoomkern bij elkaar gehouden door de sterke atoomkracht ; zo worden de electronen bij de atoomkernen bijeengehouden door electrische aantrekkingskracht, zo worden de watermolecules in een regendruppel (die bij de vorming bij benadering bolvormig is) bijeengehouden door de (zelf)gravitatie van de watermolecules en zo worden de planeten in een (bij benadering) cirkelvormige baan rond de zon gehouden.
De bolvormige symmetrie die door de Grieken zo werd aanbeden, is dus te wijten aan een intrinsiek kenmerk van bepaalde universele Alfa-attractoren, namelijk het feit dat het punt-attractoren zijn.
- Sub-Optimale symmetriëen van Alfa-organisaties
Hoewel de universele Alfa-attractoren zoals in het bovenstaande voorbeeld als puntattractoren te beschouwen zijn, zien we toch vele evolutiepatronen die afwijken van de optimale bolvorm of de cirkelvormige banen rond een puntattractor. In de meeste gevallen zijn hierbij volgende oorzaken te onderkennen
- er is niet 1 puntattractor aan het werk, maar er is een samenspel van attractoren
- de begincondities zorgen voor afwijkingen in de symmetrie van de interactor (er zijn bijvoorbeeld hindernissen of obstakels die de normale ontwikkeling van de interactor belemmeren of beïnvloeden).Ik zal hiervan enkele voorbeelden geven, om de gedachten te vestigen.
1) De aarde is niet rond, doch sferoïdaal
Newton voorspelde, precies aan de hand van zijn gravitatietheorie, dat de aarde niet rond doch licht afgeplat zou zijn aan de polen. De oorzaak voor de afplatting is dat de aarde ronddraait rond zijn eigen as, waardoor een middelpuntvliedende kracht ontstaat. Een evenwicht van krachten, met een minimale potentiële energie, bestaat in dit geval niet bij de bolvorm, doch bij een afgeplatte bol, waarvan de afplatting bepaald wordt door de snelheid van het lichaam. De middelpuntvliedende kracht is hier dus als extra kracht in het spel .
2) De baan van planeten rond de aarde is niet cirkelvormig, doch ellipsoïdaal
Het was de Duitse sterrenkundige Johannes Kepler (1571-1630) die ontdekte dat de planeten geen cirkelvormige baan rond de aarde maakten. Uit talrijke studies van de gegevens over de bewegingen van de planeten, deed hij de - voor die tijd - revolutionaire ontdekking van de elliptische banen van de planeten. Hij slaagde erin drie mathematische wetten te formuleren, die de elliptische baan van de planeten correct beschrijft. De zon staat niet in het centrum van de ellips, doch in één van de twee brandpunten van de ellips.
Hierbij is het theoretisch niet onmogelijk dat een cirkelvormige baan zou beschreven worden : een cirkel is immers een bijzondere vorm van een ellips, waarbij de twee brandpunten samenvallen. Het is echter zeer onwaarschijnlijk, omdat het slechts één van de oneindige vele ellipsvormen is, en er geen enkele reden is waarom nu precies de meest symmetrische vorm moet gekozen worden. De enige wetmatigheid waar alle planeten aan moeten voldoen, is dat er evenwicht is tussen de centripetale (middelpuntvliedende) kracht, en de gravitatiekracht : opnieuw vinden we dus een samenspel van twee attractoren. Is de zwaartekracht groter, dan zal het lichaam worden aangetrokken naar de het centrale lichaam (dat als het niet roteert, naar de bolvorm zal streven). Is de centripetale kracht groter, dan zal het lichaam uit de baan rond het centrale lichaam ontsnappen. Alle ellipsoïdale banen zijn echter mogelijke en gelijkwaardige oplossingen voor de baan rond het centrale lichaam. Welke baan zal gekozen worden, is afhankelijk van de initiële condities, met andere woorden van de initiële informatie die het samenspel van de 2 attractoren bepaalt.
3) Atomen zijn geen bollen
Atomen zijn geen harde bollen, doch dynamische evolutiepatronen waarbij de electronen in een wolk rond de kernen draaien. De exacte vorm en snelheid van de electronen, is door toevallige factoren bepaald, in die mate dat we ze nooit exact zouden kunnen bepalen. Gemiddeld gezien kunnen we de banen echter voorstellen als een bolvormige wolk, waarbinnen de electronen zich met een bepaalde waarschijnlijkheid in bewegen. De gemiddelde symmetrie is dus bolvormig.
- Symmetrie bij samenspel van Universele en contextuele Alfa-attractoren
Wanneer niet enkel een Unviversele attractor, doch ook een contextuele attractor (lokale informatie) aan het werk is, zullen de gevormde interactoren nog meer afwijken van de ideale (universele) symmetrievorm. De symmetrie zal dan lager zijn, doch wel afgeleid zijn van dezelfde "symmetriegroep".
Een mooi voorbeeld vinden we terug bij zeepbellen.
Omdat deze zo licht zijn, is hun vorm nauwelijks beïnvloed door de gravitatiekracht, doch wel door de intermoleculaire aantrekkingskrachten en de locale hindernissen. De plaats van elke molecule in de zeepbel, wordt niet enkel beïnvloed door de zwaartekracht, doch ook door de vorm en intermoleculaire kracht van de directe buren. Idealiter sluit hun vorm nauw aan bij een bolvorm, omdat het gemiddelde van alle aantrekkingskrachten tussen de molecules, het zwaartepunt van de aantrekkingskracht in het midden van de bolvorm legt. Wanneer we echter de randcondities wijzigen, en "lokale informatie" of lokale obstakels toevoegen, zien we subobtimale vormen verschijnen ; het zijn vormen die - in de gegeven omstandigheden, zo goed mogelijk de bolvorm benaderen.
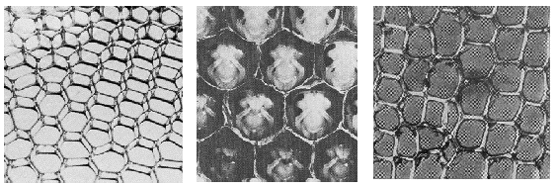
Wanneer we een reeks zeepbellen tussen 2 evenwijdige glasplaten blazen, krijgen we als resultaat een patroon van min -of meer zeshoekige cellen. Deze zeshoeken zijn nu de optimale vormen in de gegeven omstandigheden. Onderstaande figuur toont aan dat we deze zeshoekige celvormen nog op andere plaatsen in de natuur tegenkomen.
Figuur H10F3 : 6-hoekige symmetriepatronen als afgeleide van de cirkelvorm of bolvorm komen veel voor in de natuur
Links) zeshoekige cellen in zeepvliezen
Midden ) zeshoekige cellen in een honingraat
Rechts) zeshoekige cellen in een plantenweefsel
Zeshoekige patronen komen heel veel voor in de natuur. Naast bovenstaande voorbeelden kunnen we nog de zeshoekige Bénard-patronen vermelden, zeshoekige patronen in de celwand van bepaalde raderdiertjes, zeshoekige patronen in de vliezen van bepaalde insectenvleugels en talrijke zeshoekige patronen in kristallen.De reden is dat de dichte pakking van bollen in een zeshoekige cel een optimale vorm is.
Op dezelfde wijze rangschikken zich bollen tussen twee platen volgens een zeshoekig kristalrooster. De zeshoekige vorm is hierbij optimaal in vergelijking met bijvoorbeeld vierhoekige stapeling, omdat het de "dichtste stapeling", waarbij de de ruimte tussen de bollen minimaal is. Het is dus de meest efficiënte wijze om bollen te stapelen, met andere woorden binnen de gegeven omstandigheden, is dit de beste wijze.
Een andere interessante vorm die we kunnen maken met zeepbellen is de "wormvorm". Wanneer we een reeks zeepbellen aanmaken, op een dergelijke manier dat ze aan elkaar vast blijven hangen, krijgen we een "worm" van zeepbellen. Deze worm geeft de optimale vorm weer voor een reeks bollen die aan elkaar zijn gehecht : de worm is optimaal in die mate dat een maximaal volume wordt omsloten met een minimale oppervlakte. Op deze wijze is de natuur "zuinig" met zijn bouwstenen. Door middel van de variatierekening kan bevestigd worden dat dit inderdaad optimale vormen zijn. Deze wormvorm treffen we op verschillende plaatsen in de natuur aan. In de eerste plaats bij wormen of rupsen, doch ook de basisvorm van insecten (met de kop, de romp en het achterlijf) is te beschouwen als een afgeleide van een 3 ledige worm. Ook sommige bacteriën vormen kolonies die dergelijke wormvormen aannemen.
Terug naar de index van dit hoofdstuk
3.2. Symmetrie en Beta-informatie
Daar waar de universele attractoren en Alfa-attractoren resulteren in interactoren die een evenwicht bereiken, dat gekenmerkt wordt door een minimale potentiële energie en een bijpassende vorm, is dit niet het geval voor de Beta-attractoren.
Beta-interactoren danken hun vorm immers aan proces-informatie, die bestaat zolang er een niet-evenwichtsdynamiek werkzaam is.
Tijdens de fase van "quasi-evenwicht", zijn de vorm en de symmetrie aangepast aan het bekomen van een maximale flux.
- Bénard cellen
Een voorbeeld hiervan hebben we reeds besproken bij de zogenaamde "Bénard-cellen". De Bénard-cellen onstaan omdat ze overeenkomen met een maximale flux : de langwerpige cellen oefenen immers een functie van "snelweg" uit. De best passende vorm voor deze cellen is opnieuw de zeshoekige vorm. De zeshoekige vorm impliceert immers een maximale densiteit van stapeling, of met andere woorden, een maximaal aantal "snelwegen" voor een bepaalde oppervlakte.
- Golven als symmetriekampioenen
Uitstekende voorbeelden van symmetrische Beta-processen zijn golven.
Golven ontstaan ten gevolge van een lokale energiegradiënt.
Als we een steen in het water gooien, ontstaan watergolven vanuit het "stress-centrum" waar we de steen hebben gegooid. Als we in onze handen klappen, onstaan geluidsgolven die zich door de lucht voortplanten.
Beide types golven zijn gradiëntdissipatoren, die zullen blijven bestaan, tot de energiegradiënt verdwenen is.Kenmerkend voor alle golven is dat hun hoge symmetrie bepaald is door slechts 2 parameters, de golflengte en de frequentie van de golven. Deze symmetrie is de oorzaak van een aantal kenmerkende eigenschappen van golven zoals de resonantie, diffractie, enzovoort.
De reden waarom het zo efficiënte energie-gradiëntdissipatoren zijn, is dat ze energie verplaatsen, zonder de materie (volledig) mee te verplaatsen : ze verspillen dus als het ware zo weinig mogelijk energie in de materie-transfer, ten gunste van de energietransfer. De materiële deeltes bewegen lokaal, doch niet met de ganse golf mee.
- Symmetrie-patronen in de biologie
We weten dat Beta-interactoren slechts stabiel zijn binnen een bepaald dynamisch gebied. Wordt een kritische parameter overschreden, dan wordt het evolutiepatroon instabiel, en ondergaat een wijziging naar een nieuw stabiel regime. We hebben reeds eerder gezien dat bij Beta-organisaties, de vorm van de interactor volledig is afgestemd op, of in harmonie is met de functie en de dynamiek van het zelforganiserend proces : vorm en functie zijn hier noodzakelijkerwijze in overeenkomst. Zoniet worden de patronen instabiel en verdwijnen tot een patroon met een andere dynamiek en vorm.
Toch kunnen we ook bij dynamische processen, en meer bepaald bij het verbreken van symmetrieën bepaalde tendensen waarnemen, die erop wijzen dat ook hier het aantal mogelijke oplossingen voor (quasi-)stabiele vormen beperkt is. Ook bij het verbreken van symmetrieën bij zelforganiserende processen, is er een zekere vorm van "convergentie" waar te nemen.Tijdens het ontstaan van Gamma-interactoren zijn zelforganiserende processen vaak de basis geweest voor de evolutie. Doordat de ontwikkeling van het individu in zekere mate een herhaling is van de ontwikkeling van de ganse soort (Haeckel), moeten we het voorkomen van deze spontane zelforganiserende processen kunnen herkennen aan de reeksen van opeenvolgende symmetriebrekingen in de ontwikkeling van het individu. Het zoeken naar deze reeksen van opeenvolgende symmetriebrekingen in de levende systemen, is als het zoeken naar de handtekening van de meester-architect die uitblinkt in het gebruik van zelforganiserende processen. Deze architect is soms een beetje "lui", en laat de spontane processen evolueren volgens hun eigen dynamiek, zonder alles tot in de puntjes zelf te willen ontwerpen. Hij gebruikt Beta-attractoren als basismodules om zijn ontwerpen op te bouwen.
We zullen hiervan een aantal voorbeelden geven. Het is aan de lezer om in het dagelijks leven additionele voorbeelden te vinden.
Laten we bijvoorbeeld opnieuw de bol en de cirkel als optimale symmetrieën beschouwen, en laten we eens nagaan in welke reeksen hun symmetrieën kunnen worden gebroken. Daarna zullen we nagaan waar we dergelijke patronen van symmetriebreking bij levende systemen aantreffen.
Beschouwen we bijvoorbeeld een druppel olie, die in een andere vloeistof met dezelfde densiteit wordt gebracht. Veronderstel dat we de druppel met een cirkelvormige schijf raken en dat we de schijf lanzaam rond zijn eigen as beginnen draaien.
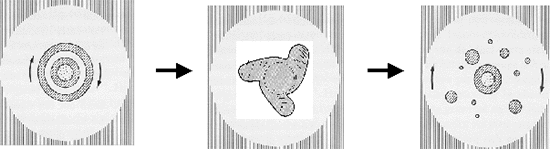
Figuur H10F4 : symmetriebreking bij een druppel olie
De druppel zal eerst platter worden (cfr. de afgeplatte sferoïde). Naarmate we echter de rotatiesnelheid van onze druppel verhogen, zullen we opeenvolgende symmetriebrekingen zien, die leiden tot opeenvolgende (quasi-)stabiele evolutiepatronen.
Eerst zullen we een ringvormige uitstulping vaststellen rond de buitenkant van de ring (fig F10F4, links). Bij snellere rotatie onstaan lobben op de ring (één tot meerlobbig, afhankelijk van de snelheid, (Fig H10F4 midden), en bij nog hogere snelheid worden satelietdruppels die zich afsplitsen van de hoofddruppel (Fig H10F4 rechts).
De opeenvolgende patronen van symmetriebreking zijn dus (in een vlakke doorsnede) :
cirkelvorming -> cirkel met ringen -> cirkel met lobben -> cirkel met satelietcirkeltjes
Elk van deze vorm heeft een lagere symmetrie dan de voorgaande vorm, en is aangepast aan de dynamiek van de interactor (hier de omtreksnelheid van de druppel). Naargelang het opdrijven van de snelheid sneller of trager gaat, kunnen eventueel tussenstappen in de symmetriebreking worden overgeslaan.In onderstaande figuur is aangetoond dat dergelijke patronen van symmetriebreking veel in de natuur rondomons kunnen worden waargenomen. Hoe verschillend de onderliggende drijfveren en interactoren ook kunnen zijn, de gradiëntdissipatie gebeurt volgens gelijkaardige symmetriebrekingspatronen.
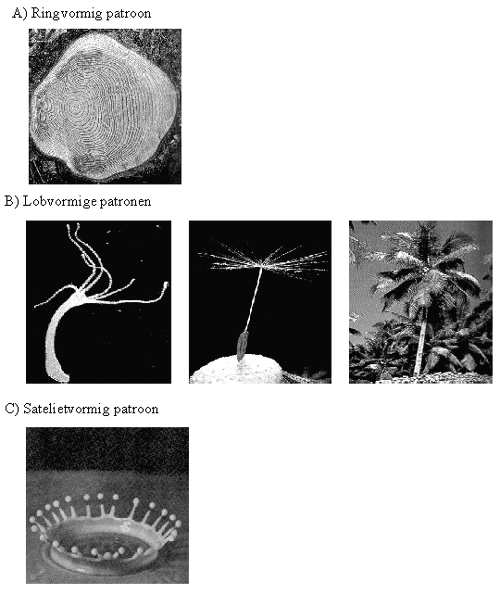
Figuur H10F5 : ringvormen, lobvormen en satelietvormen als voorbeelden van gebroken symmetrie :
a) de ringvorm bij dwarse doorsnede van een boomstam
b) lobvormige tentakels bij de zoetwaterpoliep Hydra, een pluimpje van een paardebloem en een palmboom
c) Satelietdruppels bij een vloeistofspatDe ringvormige symmetrie vinden we bijvoorbeeld terug bij dwarse doorsnede van de stengel van een plant of een boom.
Gedurende de ontwikkeling van een plant of een boom of een plant, onstaan op een bepaald ogenblik vertakkingen, net zoals er lobben ontstaan in de druppel. Op dit moment wordt dus de cirkelsymmetrie verder gebroken. De cirkelsymmetrie wordt gebroken in een M-voudige rotatiesymmetrie, waarbij de M afhangt van het type plant.Als u voortaan nog naar planten en bomen kijkt, of jonge met oude exemplaren vergelijkt, moet u maar eens letten op deze patronen van symmetriebreking.
Dezelfde patronen vinden we terug bij bijvoorbeeld de zoetwaterpoliep. Een jonge poliep is aanvankelijk een cirkelvormige cilinder, doch op een bepaald moment ontstaan vertakkingen, en wordt de cirkelsymmetrie gebroken.
Zowel bij de poliep als bij bomen en varens, kan dit proces zich eindeloos herhalen : elke vertakking kan verder gedurende een bepaalde tijd groeien, om daarna opnieuw te vertakken. Op deze wijze ontstaat de typische boomstructuur of varenstructuur.
De vorming van "satelietstructuren" als verdere vorm van symmetriebreking vinden wij bijvoorbeeld terug wanneer we een voorwerp in water of een andere vloeistof laten vallen. Eerst ontstaan de cirkelvorming golven rond de plaats waar het voorwerp is gevallen, daarna vormen zich lobben rond deze golven (waardoor de vloeistof het uitzicht van een "kroontje" krijgt), en uiteindelijk komen satelietdruppeltjes los uit de lobben van het kroontje. De drie stadia van de symmetriebreking zijn dus merkbaar bij dit mooie Beta-proces.Hoewel een draaiiende druppel en een groeiende boom of zoetwaterpoliep totaal verschillende evolutiepatronen zijn, hebben ze toch een belangrijk symmetriekenmerk gemeenschappelijk. Omdat een cirkelsymmetrie maar op een beperkt aantal wijzen kan verbroken worden, vertonen de patronen van symmetriebreking van de oliedruppel, de zoetwaterpoliep, een palmboom en een spat analoge eigenschappen. Ze ondergaan een convergente evolutie, echter nu onder invloed van een zelforganiserend proces.
- Biologische morfogenese en symmetrie
Bij de biologische morfogenese kunnen we in dit kader twee belangrijke vragen stellen.
Een eerste vraag is waarom de natuur deze zelforganiserende principes gebruikt gedurende de evolutie.
Een tweede (en moeilijkere vraag) is hoe ze dit doet, in complexe interactoren als planten en dieren.
We zullen op beiden vragen een beknopt en principieel antwoord proberen geven.
Waarom volgt de evolutie van Gamma-attractoren zelforganiserende principes en patronen van opeenvolgende symmetriebreking ?
Eerst en vooral maken Gamma-attractoren deel uit van het universum in zijn geheel, en zijn ze dus ook onderworpen aan de eigenheden (wetmatigheden) die gelden voor Alfa-atractoren en Beta-attractoren. Een koe valt net zo goed naar beneden onder invloed van de gravitatiekracht als een steen...
Ten tweede is er een belangrijk economisch voordeel aan het gebruik van deze zelforganiserende processen.
Gamma-replicatoren bevatten een enorme hoeveelheid Gamma-informatie, die de complexe ontwikkeling van de interactor in goede banen moet leiden. Door bepaalde delen van het ontwikkelingsproces over te laten aan de spontane dynamiek van zelforganiserende processen, moet voor deze fasen van de ontwikkeling geen Gamma-informatie (bijvoorbeeld genetische informatie) opgeslaan worden. In de zelforganiserende fasen van de ontwikkeling, gebeurt de ontwikkeling bij wijze van spreken "met een automatische piloot". Deze automatische piloot is een zelforganiserend proces. Dat de informatie door het gebruik van zelforganiserende processen sterk wordt beperkt, kan gemakkelijk worden ingezien aan de hand van fractaalstructuur (of zelfgelijkvormige structuur). Een varen is hiervan een mooi voorbeeld. Elke tak van de varen, wordt op een bepaalde plaats in 2 (of meer) vertakt : deze takken worden op zich verder in vertakt. Een instructie om dit proces te beschrijven kan dus als volgt luiden : "vertak na een lengte x in 2 ; herhaal dit proces telkens opnieuw". De informatie moet dus niet telkens opnieuw herhaald worden. We hebben reeds eerder aangetoond dat fractaalstructuren vaak ook wiskundig éénvoudig te beschrijven zijn. Daarenboven is de kans op het maken van fouten bij het copiëren van een éénvoudige instructie, veel kleiner dan bij het copiëren van een complexe instructie.
Bovendien hebben zelforganiserende processen de intrinsieke eigenschap quasi-stabiele stadia te vertonen. Deze stabiliteit is belangrijk voor de overleving : als alle deelprocessen van een levend systeem stabiel zijn, is de kans dat het geheel stabiel is groter...
De éénvoud, de symmetrie(breking) en de stabiliteit van bijvoorbeeld de vorm van een varen, zijn uitingen van éénzelfde aspect : de natuur heeft tijdens de ontwikkeling gebruik gemaakt van een zelforganiserende dynamiek. De Beta-informatie en de Gamma-informatie werken synergistisch, waardoor de Gamma-replicator niet alle ontwikkelingsinformatie moet bevatten.
Een tweede vraag die moet beantwoord worden, is hoe de natuur erin slaagt deze zelforganiserende dynamiek op te nemen in de evolutie van een levend systeem.
Deze brengt ons terug naar het moeilijke terrein van de morfogenese, of (letterlijk) "het onstaan van de vormen" in de biologie (we hebben dit al aangehaald bij de paragraaf over specialisatie). De eerste persoon die hiervoor een afdoende verklaring gaf is Alan Turing. Merkwaardig genoeg is dit een wiskundige, die bovendien bekendheid heeft verworven met de bouw van één van de eerste computers in 1943. Turing was een bijzondere persoon, nogal een eenzaat, met een heel breed interessegebied. In 1952 schrijft hij een artikel getiteld "The chemical basis of morphogenesis". Hierin poneert hij de indrukwekkende stelling dat een systeem van chemische stoffen, de "morfogenen" genaamd, die op elkaar reageren en door de weefsels diffunderen, voldoende is om de belangrijkste verschijnselen van de morfogenese te kunnen beschrijven.
Hij was hiermee de voorloper van Conrad Waddington, die het begrip "epigenetisch landschappen" lanceerde. Een epigenetisch landschap is functioneel te vergelijken met een potentiaallandschap. In het epigenetische landschap komen de pieken en dalen echter niet overeen met bijvoorbeeld de hoogte in meters in een reeël landschap, doch wel met de concentraties van morfogenen in een biologische weefsel. Daar waar een potentiaallandschap te beschouwen is als een semiotisch Alfa-landschap, is een epigenetisch landschap te beschouwen als een semiotisch Beta-landschap. Daar waar er pieken zijn in het epigenetische landschap, zijn er dus hoge concentraties van morfogenen. Op deze plaatsen kan de celdeling in het weefsel sterk worden gestimuleerd, wat bijvoorbeeld kan leiden tot het onstaan van een extra vertakking. Omgekeerd kan een morfogene stof de celdeling nadelig beïnvloeden, waardoor enkel ver verwijderde cellen beter delen. In elk geval zal de groei en de celdeling in het weefsel beïnvloed worden door de pieken en dalen in het epigenetisch landschap. De pieken en dalen in het epigenetisch landschap zelf, ontstaan door een zelforganiserende dynamiek, die de morfogenen in golven laat onstaan en/of diffunderen. In dit geval was misschien beter gesproken van een "epigenetische oceaan", waarbij de hoogte van de golven overeenkomt met de concentratie aan morfogenen. Turing merkte reeds op dat verschillende types golven (staande of rollende), tot een verschillende morfogenese kunnen leiden bij plant en dier. In een cilindervormig weefsel leiden de "rollende golven" tot een patroon waarbij de morfogenen zich in ringen rond de centrale as verspreiden (zoals bij het vallen van een steen in het water, de golven zich in kringen rond de steen verspreiden). Het gevolg is dat het weefsel opeenvolgend meer (of sneller) en minder gaat groeien. Deze rollende golven kunnen dus bijvoorbeeld de ringen in een doorsnede van een boom verklaren (figuur H10F5). Naast de rollende golven zijn er ook de "staande golven", die in een cilindervormig patroon gaan leiden tot hoge concentraties op bepaalde plaatsen van de omtrek. Deze dynamiek van de staande golven kan de verbreking van de cirkelsymmetrie in een M-voudige rotatiesymmetrie verklaren (figuur H10F5). Dit type symmetriebreking kan dus de vorming van takken in de stam van een boom verklaren, de vorming van bloemblaadjes in een bloem, of de vorming van tentakels in een poliep of een zee-anemoon.
Wanneer we terugkomen op het voorbeeld van celdifferentiatie bij de kikker (zie vorig hoofdstuk), zien we dat ook deze ontwikkeling gedeeltelijk bepaald is door een zelforganiserende dynamiek. Ook hier verraadt de architect immers zijn werkmethodes.
De aanvankelijke sferische symmetrie van de bevruchte eicel, wordt eerst gebroken door een tweedeling in twee symmetrische helften : hierbij is de bolsymmetrie gebroken in een spiegelsymmetrie. Door de opeenvolgende celdelingen, wordt de spiegelsymmetrie verder gebroken. Tot hier dus een eerste fase van opeenvolgende symmetriebrekingen. Uiteindelijk krijgen we het kiemblaasje, dat opnieuw een sferische symmetrie heeft (de symmetrie is wel lager dan de symmetrie van de bevruchte eicel). Tijdens de gastrulatie wordt de sferische symmetrie van het kiemblaasje, verbroken tot een cilindervormige symmetrie : de gastrula is bij grove benadering te beschouwen als een dikke buis. In deze fase is de sferische symmetrie dus verbroken in een cilindersymmetrische symmetrie. In de volgende fase, de neurulatie, wordt deze "buis" min of meer platgedrukt, en ontstaan uitstulpingen, waardoor de cilindersymmetrie wordt gebroken in ruwweg een spiegelsymmetrie. Deze uitwendige spiegelsymmetrie zal gedurende de ganse ontwikkeling van het organisme behouden worden (ledematen, ogen en dergelijke verschijnen symmetrisch aan elke kant van het lichaam).
Deze opeenvolgende symmetriebrekingen wijzen er opnieuw op dat de ontwikkeling van het embryo gedeeltelijk het gevolg is van een zelforganiserende dynamiek. Het DNA bevat dus niet alle informatie voor de ontwikkeling, doch net voldoende om het ontwikkelingsproces in goede banen te leiden. Wanneer we naar een andere stad rijden met de wagen, hebben we ook niet een uiterst gedetailleerde wegbeschrijving nodig, die ons elke millimeter van de weg, tot in de kleinste details beschrijft. Wat we nodig hebben is een plan, waar de cruciale beslissingspunten aangeduid zijn door wegwijzers (selectie-informatie) : buiten deze beslissingspunten volgen we spontaan de kronkels van de weg. Op gelijkaardige wijze bevat het DNA de wegwijzers voor de ontwikkeling, doch niet een gedetailleerde beschrijving van de paden. Dus niet : cel 53210 moet op coordinaten x,y,z worden geplaatst op tijdstip a. De details van de ontwikkeling worden bepaald door de zelforganiserende dynamiek, die de paden vormt in de epigenetische landschappen. De grote lijnen van de ontwikkeling worden gestuurd door het DNA. Alfa-, Beta- en Gamma-informatie gaan dus hand in hand bij de ontwikkeling van levende systemen.
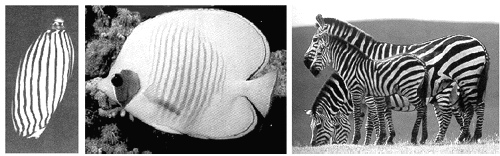
Dat deze theorieën juist zijn kan geverifieerd worden met computerexperimenten voor heel specifieke gevallen. Een prachtig voorbeeld vinden we bij de patronen van een een schelpdier "Bankivia Fasciata" dat we kunnen aantreffen in het Groot-Barrière rif bij Australië (onderstaande figuur links). De onderzoekers Meinhardt en Klinger slaagden erin een mathematisch model te ontwikkelen, waarbij ze de reacties en diffusie van morphogenen trachten te voorpellen. Hun voorspelling wordt gevisualiseerd door middel van computerbeelden.
Figuur H10F6 : een uitwendig patroon bij een schelp, een tropische zeevis (Chaetodon) en een zebra
Op dezelfde wijze kan wellicht het ontstaan van complexe uitwendig patronen worden verklaard zoals bijvoorbeeld de strepen op de zebras en de vlekken van luipaarden.
Analoge computersimulaties zijn uitgevoerd om bijvoorbeeld het ontstaan van wortels in bepaalde plaatsen van planten te simuleren. Ook een volkomen ander type proces, de ontwikkeling van compartimenten in de vleugels van vliegen, kan men met dergelijke mathematische modellen simuleren. Ook hier slaagt men erin modellen te bouwen die een resultaat hebben dat de werkelijkheid zeer dicht benaderd, wat er opnieuw op wijst dat de onderliggende principes wellicht correct zijn.De werkelijkheid is echter nog veel complexer dan hierboven geschetst. Tal van ontwikkelingsmechanismen kunnen gezamelijk de interactiepatronen mede beïnvloeden. Zo is een bevruchte eicel aanvankelijk niet volledig homogeen, en worden de eerste klievingen beïnvloed door de niet homogene concentratieverdeling van bepaalde chemische substanties. Dit komt er dus op neer dat bepaalde cellen, onder invloed van concentraties van chemische stoffen, tot specifieke interactoren zullen leiden (zoals bijvoorbeeld vingers, trilharen, enz...).
Een ander belangrijk concept in de morfogenese is dat van de "positionele informatie". Deze theorie stelt dat de cel gedurende zijn ontwikkeling continu zijn "positie" bepaalt in het ontwikkelend weefsel, en dat het deze positie is die de uiteindelijke toekomst van de cel zal bepalen.
Deze mechanismen maken deel uit van wat eerder de "interpretatie" van de Gamma-informatie werd genoemd.Terug naar de index van dit hoofdstuk
3.3. Symmetrie en Gamma-informatie
3.3.1. Natuurlijke selectie en optimale vormen
Eén van de basisprincipes van de natuurlijke selectie, is dat in de natuur toevallige variaties ontstaan, waarvan de beste varianten een competitief voordeel hebben ten opzichte van de minder optimale varianten. Als gevolg hiervan onstaat een drift in de genenpool, die leidt tot de ontwikkeling van levende systemen die alsmaar beter zijn aangepast aan hun omgeving. Natuurlijke selectie is dus een optimaliserend principe dat - zij het op zeer langzame wijze - tot opzienbarende resultaten kan leiden. Deze optimalisatie uit zich ook in een toenemende symmetrie.
Een heel mooi voorbeeld van convergentie evolutie in dit verband is de symmetrie van snelzwemmende zeedieren.Beschouwen we bijvoorbeeld een dolfijn, een haai en een pinguin. Deze 3 snelzwemmende waterdieren hebben allen een vorm die kenmerkend is voor het leven van snelbewegende dieren in water. Eerst en vooral vertonen ze een spiegelsymmetrie, met vooraan de uitwendige sensoren, zoals dit ook bij andere land- en zeedieren is die zich relatief snel voortbewegen. Bovendien hebben ze alle drie vinnen, gebruikt voor de voortstuwing en de sturing en ze hebben een sterk gestroomlijnd lichaam, met een vrij spitse snuit. Nochtans is de dolfijn een zoogdier, dat afstamt van de landdieren, en is de pinguin een afstammeling van de vogels. Deze gelijklopende ontwikkeling van de uitwendige vorm is te verklaren door het feit dat voor een snelle lineaire voortbeweging in het water, deze gestroomlijnde vorm, en de wijze van voortstuwing, optimaal is. Wanneer we de dieren echter bekijken vanuit anatomisch standpunt, zullen we zien dat onder het schijnbaar gelijkaardige uiterlijk, er belangrijke onderhuidse verschillen zijn. Zo zijn de borstvinnen van de pinguin afgeleid van de vleugels van de oorspronkelijke vogel, en zijn de borstvinnen bij de dolfijn afgeleid van de voorpoot van een vleesetend zoogdier. Voor eenzelfde probleem in de evolutie, zijn maar een beperkt aantal optimale oplossingen, waardoor de vormen van deze waterdieren naar elkaar toe zijn gegroeid.
Terug naar de index van dit hoofdstuk
3.3.2. Artificiële constructies en symmetrie
Artificiële constructies zijn steeds door de mens ontwikkeld met een bepaald doel voor ogen. Alle artefecten voldoen in zekere mate aan een bepaalde behoefte die leeft bij een groep mensen. Deze groep noemt men in marketingtermen "de doelgroep". Commerciële bedrijven doen wetenschappelijke studies van deze doelgroepen en hun specifieke eisen. Zo zal men bij de ontwikkeling van een nieuwe wagen, een boot of een vliegtuig, de specifieke vereisten van de doelgroep gaan vertalen in "productspecificaties", die nauwkeurig alle kenmerken van het te bouwen artefact beschrijven. Deze productspecificaties worden in het bedrijf vertaald in processpecificaties, die het geheel van processen in de onderneming zodanig beschrijven, dat ze met een optimaal evenwicht van kosten versus toegevoegde waarde, aan de wensen van de klant voldoet.
Ook de mens is echter bij zijn artificiële constructies beperkt in het aantal mogelijke constructies voor een bepaalde functie of proces. Het is dan ook niet verwonderlijk dat bijvoorbeeld een duikboot, min of meer dezelfde aerodynamische vorm heeft als een vis, of dat een vliegtuig dezelfde basisvorm heeft van een vogel. Ook de symmetrie van deze voorwerpen is convergent met de snelbewegende biologische levende systemen.
In beide gevallen heeft de mens echter een ander principe gebruikt voor de voortbeweging. In het geval van de duikboot gebeurt de voorstuwing door een schroef. De natuur heeft het principe van een roterende schroef niet "geselecteerd" tijdens de natuurlijke selectie. Hetzelfde kan gesteld worden voor de schroeven of turbines van een vliegtuig. Schroeven (voor beweging in een medium) en wielen (voor beweging op een medium) zijn uitstekende middelen om rotaties om te zetten in translaties, en het is enigszins merkwaardig dat biologische wezens deze niet als natuurlijke subsystemen hebben ontwikkeld. Dit toont alleen maar aan dat natuurlijke selectie leidt tot het selecteren van meer en meer optimale vormen, zonder dat dit daarom de meest optimale vorm hoeft te zijn.Terug naar de index van dit hoofdstuk
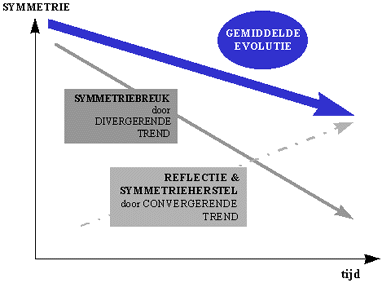
3.4. Symmetrie en symmetriebreking in de evolutie
De voorgaande paragrafen kunnen we als volgt samenvatten :
Aanvankelijk was er in ons heelal weinig verspreiding, en dus een hoge symmetrie.
Door de toenemende verspreiding is echter ook de symmetriebreuk steeds toegenomen.
Conform met het principe van de fluxmaximalisatie, wordt ook de symmetriebreuk of "verspreiding van symmetrieën" enigszins beperkt. Dit is onder twee vormen zichtbaar.
Enerzijds is het zo dat, bij een symmetriebreuk vanuit een interactor met een hoge symmetrie, de nieuwe symmetrie nog steeds bepaalde kenmerken van de oorspronkelijke symmetrie zal vertonen. De symmetriebreuk verloopt dus veelal in sequenties die tot dezelfde symmetriegroep behoren.
Ten tweede zorgt het principe van de zelforganisatie en selectie voor de evolutie naar meer optimale vormen, en dus ook optimale symmetrieën voor bepaalde evolutionaire problemen. Daardoor is er een convergentie van symmetrieën waar te nemen. Door deze convergentie evolueren van oorsprong verschillende interactoren, op analoge symmetrische wijze.Ook op het vlak van symmetrie balanceert de gemiddelde evolutie dus op een lijn tussen symmetrie en assymetrie.
Terug naar de index van dit hoofdstuk
4. Integratie versus differentiatie in de evolutie
In het vorige hoofdstuk hebben we verschillende mechanismen gezien die leiden tot differentiatie van de interactoren en attractoren.
In de gemiddelde evolutie zal een fase van differentiatie veelal gevolgd worden door een fase van integratie. De differentiatie heeft immers gezorgd voor het onstaan van nieuwe subsystemen, systemen of processen, die daarna geleidelijk aan beter gecoördineerd en geïntegreerd gaan verlopen. Om deze reden kan gesproken worden van een "differentiatie-integratie"-cyclus in de evolutie, waarbij fasen van differentiatie en integratie elkaar opvolgen.
Daar waar de fase van differentiatie eerder gekenmerkt is door een schijnbaar streven naar toenemende specialisatie en de uitbouw van nieuwe variaties op bestaande themas, is de fase van integratie meer gekenmerkt door een schijnbaar streven naar harmonizatie. Daardoor kan gesteld worden dat de differentiërende fase eerder gekenmerkt door innovatie, terwijl de integrerende fase eerder gekenmerkt is door confirmatie, met verhoogde stabiliteit tot gevolg. De differentiërende fase is eerder een fase van chaos en symmetriebreuk, terwijl de integrerende fase een eerder een fase is van herstel van de symmetrie en de orde. Bij levende systemen (zoals cellen of levende organisaties) is de immature of jeugdige fase voornamelijk gekenmerkt door differentiatie, terwijl de mature fase voornamelijk een fase is van integratie, gepaard gaande met een toenemende redundantie en controle van de stromen van materie, energie en informatie.
Hoe wezenlijk deze differentiatie en integratie de evolutie kenmerken, zullen we in de volgende paragrafen bekijken. We zullen zien dat voor een individueel systeem of proces deze fasen kunnen onderscheiden worden, doch dat in de evolutie alle systemen en processen gezamelijk interageren en evolueren, waardoor ook deze fasen van differentiatie en integratie elkaar doorkruisen.Terug naar de index van dit hoofdstuk
4.1. Differentiatie, integratie en co-evolutie
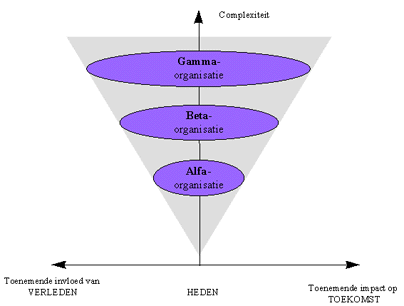
Door de toenemende specialisatie, zijn gedurende de evolutie verschillende niveaus van interactoren gevormd (bijvoorbeeld atomen, molecules, cellen, organismen, enz...).
Aangezien echter elke interactor deel uitmaakt van de omgeving van alle andere interactoren, en de evolutie een spontane trend vertoont naar aanpassing aan de omgeving, is de evolutie van elk evolutiepatroon niet los te koppelen van de andere interactoren in zijn omgeving. Dit is de basis voor het begrip "co-evolutie". Co-evolutie betekent dat de verschillende evolutiepatronen samen, op een interactieve manier evolueren, die ervoor zorgt dat ze steeds beter aan elkaar zijn aangepast. Co-evolutie is dus een aspect van integratie in de evolutie, dat leidt tot convergentie tussen de verschillende interactoren en interacties. Op verschillende niveaus in de evolutie kunnen we voorbeelden geven van integratie tussen het macroniveau en het microniveau (wat ik eerder "verticale integratie" heb genoemd).
- Co-evolutie in het universum
In een eerste fase van de evolutie (de cosmische evolutie), was de evolutie voornamelijk gekenmerkt door (Universele) Alfa-attractoren en Beta-attractoren. Wellicht zijn in de initiële hoog-energetische energiestroom eerst de eerste Beta-attractoren ontstaan. Vanaf het ontstaan van de eerste materie (en dus ook de eerste Alfa-attractoren), was er op "macro"-schaal een condensatie van materie waar te nemen tot de grote structuren in het heelal, gaande van galactische nevels tot sterrenstelsels en superclusters. Aan de "micro"-kant van de evolutie, onstonden door afkoeling meer en meer atomen en molecules. Nadien zijn nog talrijke, doch minder universele Alfa- en Beta-attractoren ontstaan, met een steeds lagere symmetrie en een steeds hogere complexiteit. Over het algemeen ("gemiddeld gezien") is de complexiteit van Beta-attractoren hoger dan dat van Alfa-attractoren, door het dynamisch karakter.
Sterren zijn interactoren waar beide samen evolueren : door de macroscopische gravitatie ontstaat een zodanig zware massa, dat deze leidt tot de vorming van een ster. In deze ster onstaan door chemische en nucleaire reacties, nieuwe atomen (microniveau). De chemische processen in de ster gaan verder in de loop der tijden, en zullen uiteindelijk de toekomst van de ster bepalen. Afhankelijk van de massa van de ster en de leeftijd, zullen de sterren uitgroeien tot witte dwergen, supernovas, pulsars of zwarte gaten. Zwarte gaten op hun beurt oefenen een heel grote invloed uit op de evolutie van de materie (zowel micro- als macro) in hun omgeving, doordat ze deze letterlijk opslorpt met zijn reusachtige zwaartekracht. Naarmate de macro-evolutie en de micro-evolutie verder vorderen, gaan ze meer en meer convergeren. Zowel de integratie als de voortdurende differentiatie verlopen onder invloed van de (Universele) Alfa-attractoren en Beta-attractoren. Naarmate de evolutie verder vordert, evolueren de "macro-processen" en de "micro-processen" steeds meer naar elkaar toe. Dit is gevisualiseerd in de onderstaand figuur.
Figuur H10F7 : coevolutie tussen micro en macro in kosmos
In een volgende fase van de evolutie, zijn levende systemen onstaan, ten gevolge van het onstaan van Gamma-attractoren. Hierdoor ontstond een nieuw type systemen, die we de levende systemen genoemd hebben. Ook hier vinden we een co-evolutie terug tussen de macrosystemen en de microsystemen.
Darwin heeft uitvoerig beschreven en uitgelegd hoe de evolutie van individuele organismen (micro-niveau) afhankelijk is van de omgeving of ecosysteem (het macro-niveau). De evolutie van het ganse ecosysteem, is echter ook afhankelijk van de ontwikkeling van de individuele soorten. Zo zijn bijvoorbeeld de koraalpoliepen verantwoordelijk voor de vorming van koraalriffen. Deze vormen soms stenen bastions van duizend kilometer lang langs tropische kusten, en geven hierbij levenskansen aan een enorme diversiteit aan mariene fauna en flora. Deze koraalpoliepen leven in symbiose met bepaalde algen (de dinoflagellaten) die door hun fotosynthese voor de nodige energietoevoer uit het zonlicht zorgen. Deze 2 partners zijn dus de basis voor het bestaan van de uiterst complexe en delicate ecosystemen die de koraalriffen vertegenwoordigen. Dit is een voorbeeld van "horizontale integratie", of integratie binnen een evolutieniveau.
Micro- en macro niveau evolueren dus gezamelijk en interactief, en dit geldt voor de diverse types levende systemen, gaande van de eenvoudige ééncelligen in de evolutionaire oersoep, tot de complexe organismen in een moderne maatschappij. Door deze co-evolutie is er een voortdurende reflectie van de eigenschappen van het macro-niveau in het micro-niveau en omgekeerd. Hierdoor gaat de evolutie van de verschillende niveaus convergeren.
- Co-evolutie in ecosystemen
Deze verwevenheid van micro- en macro-niveau in de evolutie geldt niet enkel voor de uitwisseling van informatie, maar ook voor de stromen van energie en materie.Een mooi voorbeeld van de verwevenheid van micro- en macro-energiestromen, en de integratie tussen de evolutie en het leven van individuen, krijgen we wanneer we de voedselketen in een ecosysteem beschouwen.
Over het algemeen kan gesteld worden dat het leven in een ecosysteem sterk bepaald wordt door de beschikbaarheid van energiestromen. Dit werd voor het eerst vastgesteld door Charles Elton, een jonge bioloog van de Oxford University. Elton begon de ecologie te bestuderen van een eiland in het Noorden van Noorwegen, rond 1920. Hij stelde vast dat er een verband bestond tussen het aantal planten, insecten en vogels en andere dieren op het eiland. Alle levende wezens maken deel uit van één continue energiestroom op het eiland, die hij de "voedselketen" noemde. In een eerste stap van de voedselketen wordt het zonlicht (macro-niveau) door de planten (micro-niveau) verwerkt via de fotosynthese, en na conversie omgezet in chemische energie. De opgeslagen energie wordt dan verder verwerkt door een plantenetend dier (een herbivoor), dat op zijn beurt wordt opgegeten door een vleesetend dier (of carnivoor). Het was Elton die binnen de biologische gemeenschappen niveaus van energietransfer begon te definiëren. Het laagste niveau wordt gevormd door de planten en de andere fotosynthetische organismen. Dit zijn de zogenaamde "producenten". Elton catalogeerde hiertoe op zijn eiland de bijvoorbeeld de grassen, bladeren van de bomen, vruchten, bloemen en zeeplanten. De zonne-energie die ze opslaan onder de vorm van koolhydraten en andere componenten wordt dan verder verwerkt door een aantal opeenvolgende niveaus van consumenten. Een insect dat een bes versnapert is een primaire consument, terwijl de vogel die het insect opeet een secondaire consument is. De vogel kan op zijn beurt opgegeten worden door een vos enzovoort... Op het einde van de voedselketen, en ook op elk niveau, vinden we de "afbrekers" terug en de "reducenten". De afbrekers zijn voornamelijk bacteriën en schimmels, die de restanten van het organisch materiaal afbreken, terwijl de reducenten zich voeden met het afgebroken organische materiaal. Op deze wijze is de voedselketen rond, want de afgebroken materialen zijn opnieuw beschikbaar voor de producenten, voor de groei van bestaand of de aanmaak van nieuw biologisch leven. Op deze wijze maakt elk levend wezen deel uit van een complexe "voedselpiramide".
Elton merkte immers op dat het aantal planten (de producenten), groter is dan het aantal primaire consumenten, dat op zijn beurt kleiner is dan het aantal secundaire consumenten (bijvoorbeeld kleine carnivoren), dat op zijn beurt kleiner is dan het aantal grote carnivoren. Hoe hoger we in deze voedselpiramide kijken, hoe groter de dieren die we meestal waarnemen. Als we alle dieren in een bepaalde categorie samennemen, en we rekenen dit om in biomassa of energie (het aantal calorieën of Joules dat elke groep vertegenwoordigt), dan krijgen we gewoonlijk een piramide van de biomassa. De grootte van de piramide reflecteert de energieflux in het ecosysteem, die in evenwicht is met de draagkracht van de omgeving.
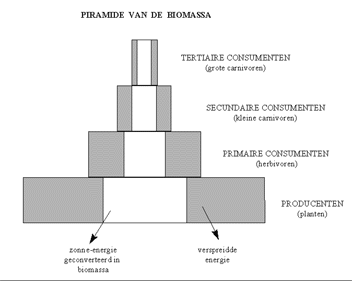
H10F8 : De transfer van (zonne-)energie van één niveau naar een ander niveau is nooit 100% efficiënt, waardoor de biomassa in elk niveau geringer is.
Zoals de tweede wet van de thermodynamica voorspelt, is de transfer van energie van het ene niveau naar het andere niveau nooit 100% efficiënt. In elk van de ontelbare biochemische reacties en fysische activiteiten die een levend systeem kenmerken, is er altijd verspreiding van energie waar te nemen (bijvoorbeeld onder de vorm van warmte). Als gevolg hiervan is minder energie beschikbaar voor elk hoger niveau van de piramide, wat resulteert in een kleinere hoeveelheid biomassa. Deze biomassa vertegenwoordigt dus het (nog) niet verspreidde deel van de energie.
Plantenecologen hebben de productie van plantaardige biomassa vergeleken in diverse types ecosystemen. De conclusies hieruit bevestigen opnieuw de uitgebreide wetten van de thermodynamica en de principes van de fluxmaximalisatie. Er blijkt duidelijk dat de productie van biomassa (micro-niveau) sterk verschilt naargelang het klimaat en de regenval (macroniveau). Equatoriale koraalriffen zijn de meest productieve biologische ecosystemen op aarden, dicht opgevolgd door estuaria en tropische regenwouden. Een indrukwekkende conclusie van dergelijke ecologische studies is dat de mens ongeveer 40% verbruikt van de totale productie van biomassa op aarde. Dit is wellicht het hoogste percentage van energieconsumptie door één enkele soort in de ganse evolutie van de aarde. De mens verbruikt deze biomassa onder de vorm van landbouwgewassen en vee, de vernietiging en de uitbating van bossen en de conversie van begroeide landschappen in door de mens bewoonde gebieden, of tot woestijnen. Aangezien de menselijke populatie steeds aangroeit, zal dit verbruik steeds stijgen (de flux wordt gemaximaliseerd), en zal er minder biomassa beschikbaar zijn voor andere consumenten op aarde. In deze context is het dan ook niet verwonderlijk dat de biodiversiteit in bijvoorbeeld tropische regenwouden er drastisch op achteruit gaat. Deze daling van de biodiversiteit, door het hoge verbruik van biomassa, is een apart voorbeeld van hoe co-evolutie tussen levende systemen werkt. De enige manier om dit te beperken is om in onze wegwerpmaatschappij een idee te integreren vanuit de klassieke biologische systemen, namelijk recyclage van energie en materialen. De cyclus van productie en consumptie zou volledig moeten gesloten worden door het uitbouwen van een volledig systeem van afbraak van de geproduceerde afvalstoffen, zodat de stroom van materie en energie omgevormd wordt tot een zelfondersteunende cyclus.Co-evolutie zorgt dus voor een verregaande integratie tussen de verschillende niveaus in de evolutie. Dit is één van de oorzaken van de harmonie die we in sommige systemen waarnemen. Het dwingt ons ook om continu te beseffen dat elke indeling of classificatie in niveaus niet de ware aard is van ons universum, doch dat deze classificaties slechts een tijdelijke en zeer contextueel bepaalde waarde kunnen hebben.
Hierbij moet ook opgemerkt worden dat ook de harmonie die we soms ervaren in het universum, zoals bijvoorbeeld in mature ecosystemen als koraalriffen of tropische regenwouden, in zekere zin ook een beperkte illusie is. Wellicht gaat het er voor de levende systemen nergens wreder aan toe. De notie van de voedselpiramide leert ons immers dat elk niveau in de piramide, het verse voedsel is voor het bovenliggende niveau. De "survival of de fittest" is in wezen een "strijd om de etsen van het zonlicht", waarbij er slechts enkelen de top kunnen bereiken en een zeker gevoel van veiligheid kunnen bemachtigen.
Terug naar de index van dit hoofdstuk
4.2. Integratie van ruimte en tijd
Gedurende de evolutie is er een toenemende integratie van ruimte en tijd, wat betekent dat in het heden, meer en meer invloeden van het verleden zijn te merken, en dat in het heden meer en meer evolutiepatronen van de toekomst worden bepaald en ontworpen.
Figuur H10F9 : impact van de evolutie op het verleden, het heden en de toekomst
Naarmate de evolutie vordert, ontstaan complexere interactoren, die in het heden meer en meer het verleden en de toekomst reflecteren. De verleden en de toekomstige tijd worden meer en meer ge-integreerd in het heden.
Het meest uitgesproken voorbeeld is hier opnieuw de mens. Aangezien een mens in essentie niks meer of minder is dan een gesofisticeerde stroom van energie en materie, gestuurd door informatie, kunnen we dit aantonen door bijvoorbeeld na te gaan hoe de mens gedurende zijn evolutie met energie is omgesprongen.
Tot voor de ontdekking van het vuur door de mens, was hij volledig aangewezen op het verbruik van voedsel voor zijn energievoorziening. De mens jaagde op dit voedsel op een gesofisticeerde wijze, zodat hij terecht tot de top van de voedselketen behoorde. We hebben in één van de vorige paragrafen gezien dat de mens hedentendage een zeer groot aandeel van de totale biomassa op aarde verbruikt. Deze energieconsumptie is kenmerkend voor de impact van de mens niet enkel in het heden, doch ook in het verleden en de toekomst. Vandaar dat het interessant is om de energieconsumptie van de mens eens in zijn historisch kader te plaatsen.Wanneer de holbewoners het vuur leerden gebruiken (ongeveer 450000 jaar geleden), was dit een revolutie in hun bestaan. Door het verbranden van hout, konden ze energie gebruiken die lang voordien door de natuur spontaan was opgebouwd. Ze konden energie (materie) die in het verleden was opgebouwd, in het heden omzetten tot bruikbare energie. Het gebruik van het vuur heeft hun leven op verschillende manieren beïnvloed. Met het vuur konden ze hun voeding koken, waardoor het niet alleen gemakkelijker te eten was en beter smaakte, doch ook de kans op microbiële infecties verkleinde. Door het gebruik van vuurtoortsen, waren ze in de mogelijkheid om snachts hun weg beter te vinden en beschermde het vuur hen van dierlijke predatoren die de mens als een element van hun voedselketen beschouweden. Bovendien konden ze hun houten hulpmiddelen harder maken, door het verharden van de punten in vuur. Doordat het vuur ook hun leefomgeving kon verwarmen, konden ze in koudere gebieden gaan leven, en zich verspreiden over de ganse aarde. De integratie van materie en energie uit het verleden, in het heden, heeft de "struggle for life" van de mens gemakkelijker gemaakt, en was heel belangrijk voor het verhogen van de kans op een lang voortbestaan van zowel de individuen, als de ganse soort.
Eén van de belangrijkste natuurlijke brandstoffen was millenia lang steenkool, dat gevormd is door biomassa die miljoenen jaren geleden in de aarde werd begraven. De grote steenkoollagen in de aarde zijn gevormd uit een overvloed aan bomen, varens en andere planten . Nadat deze planten afstierven, kwamen ze bijvoorbeeld terecht in meren, waar ze langzaam afbraken tot een slijmerige colloïdale materie. Door de tectonische bewegingen op de aarde, werden de aardplaten nu eens overspoeld door de zeeën, terwijl ze in een volgende fase weer droog kwamen te liggen. Als gevolg hiervan hebben verschillende sedimentlagen zich afgezet boven de massas afgestorven planten. Door de steeds toenemende druk van deze lagen, werd deze massa droger en harder, om uiteindelijk via een aantal overgangsstadia de steenkool te vormen.Archeologen hebben ontdekt dat meer dan 3000 jaar geleden in China de verbranding van steenkool werd gebruikt voor het smelten van koper. De Grieken gebruikten de verbrandingsenergie vanaf de vierde eeuw voor Christus. Ook de Romeinen gebruikten steenkool in de diverse delen van hun imperium, waardoor het gebruik ervan werd verspreid over Europa. Door het gebruik van deze energie uit een verleden dat miljoenen jaren verwijderd is van het heden, konden deze vroege beschavingen hun voortbestaan in de toekomst bevorderen, onder andere door de productie van betere wapens en gereedschappen.
Ook nu nog wordt steenkool veel gebruikt voor het het produceren van stoom in electrische centrales. Door de omzetting in electrische energie, kan de energiestroom veel doelgerichter gebruikt worden.Het zal echter tot de twintigste eeuw duren vooraleer de ontsluiting van de massale hoeveelheden energie uit de atoomkernen kan worden gebruikt. De energie die vrijkomt bij de splitsing of fusie van atoomkernen is energie die gevormd werd in een heel vroege periode van het onstaan van het heelal, toen de quarks condenseerden tot de eerste kernen. Hoe dieper we dus in de materie doordringen voor het produceren van onze energie, hoe massaler de energiehoeveelheden zijn die we kunnen vinden, en hoe verder deze in het verleden zijn gevormd.
Het is ook echter door het vinden van deze steeds grotere energiebronnen, dat we in staat zijn om plannen te maken in het heden en deze plannen te realiseren in de toekomst. Hoe verder we in het verleden teruggrijpen voor de winning van energie, hoe massaler de energie die beschikbaar komt, en hoe meer we de toekomst naar onze hand kunnen zetten. De impact op de toekomst is zowel positief als negatief. We kunnen de massale energiehoeveelheden gebruiken voor de constructie van imposante bouwwerken en autosnelwegen, voor de creatie van raketten om de ruimte te veroveren, enz... Onze steeds toenemende mobiliteit is een teken bij uitstek van de integratie tussen ruimte en tijd. Toen Caesar de dappersten van alle Galliërs wou verslaan (de Belgen) moest hij hiervoor een wekenlange tocht ondernemen vanuit Rome. Een modern straalvliegtuig legt deze afstand af in minder dan een uur. Tijd en ruimte zijn dus zeer relatieve begrippen, niet enkel voor wie tegen de snelheid van het licht reist.
Negatief is echter dat we niet enkel passagiersvliegtuigen, doch ook gevechtsvliegtuigen kunnen produceren. De piloten maken geen gebruik van zwaarden, speren en schilden, doch kunnen atoomwapens met zich mee transporteren, om onze imponerende artefacten te vernietigen. Andere nefaste effecten uiten zich langzaam in ons milieu : door het massale gebruik van fossiele brandstoffen onstaan grote hoeveelheden CO2, wat allesbehalve positief is voor het biologisch leven.
Ook deze negatieve factoren maken dat de mens niet alleen meer en meer in zijn verleden graaft, doch ook meer en meer de toekomst in de hand heeft.
Deze evolutie heeft op zich ook een zelfversnellend effect. Door een grotere mobiliteit (van informatie, materie en energie), zowel in de tijd als in de ruimte, is de kans op innovatie en differentiatie ook groter. Op zijn beurt stijgt ook de noodzaak voor integratie van al deze informatie en kennis, van deze nieuwe materievormen en artefacten. Zo lijken er nu bijvoorbeeld meer conflicten (tussen mensen, organisaties, of landen) dan ooit te ontstaan. Deze perceptie is echter ongetwijfeld beïnvloed door de toenemende communicatie en mobiliteit. Ze maakt ook de noodzaak hoger aan overkoepelende structuren en organisaties, die moeten zorgen voor betere samenwerking en controle van deze conflicten. Met andere woorden, doordat de innovatie en differentiatie zo groot is, ontstaat ook de noodzaak voor een onmiddellijke integratie (eerder dan te wachten op een "volgende fase" in de evolutie).
Terug naar de index van dit hoofdstuk
4.3. Integratie tussen Materie/Energie en Informatie
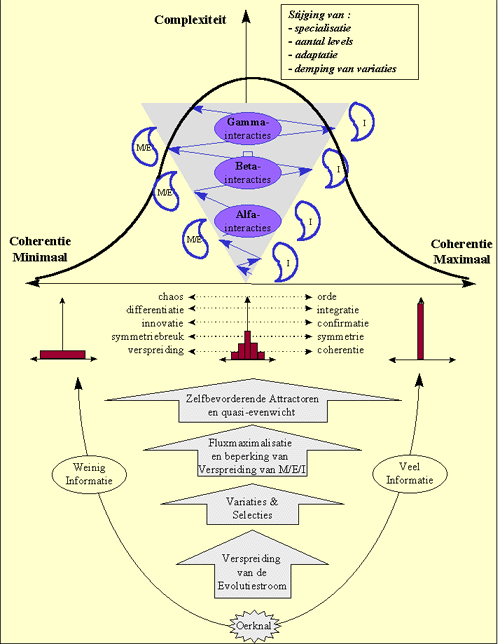
In de voorgaande hoofstukken werd reeds verduidelijkt dat de evolutie een opeenvolging is van symmetriebrekingen. De symmetriebreking waarbij een bepaald deel van materie/energie een nieuwe betekenis krijgt binnen een bepaalde context, met andere woorden het ontstaan van nieuwe informatie, werd hierbij "de meest fundamentele symmetriebreking" genoemd. Het zijn immers de diverse types informatie die ervoor zorgen de de stromen van energie en materie op geordende wijze verlopen, en zo gestalte geven aan de diverse types evolutiepatronen. We hebben hierbij het kunstmatige onderscheid gemaakt tussen Alfa-, Beta- en Gamma-informatie.
Het feit dat attractoren spontaan ontstaan en hiermee aan de basis liggen van spontaan evoluerende evolutiepatronen, noodzaakt een heel sterke integratie tussen enerzijds de informatie in de attractor en anderzijds de stroom van materie en energie. Dit hebben we aangeduid met de term "complementariteit". Doordat de materie/energie en de informatie in de attractor samen zijn ontstaan en ge-evolueerd, zijn ze volledig op elkaar afgestemd : het ene vult het andere volledig aan.Deze complementariteit geldt zowel voor Alfa-, Beta-, als Gamma-attractoren.
Nochtans is ook de relatie tussen M/E en I voor de drie types attractoren ge-evolueerd in de loop der tijden. We hebben eerder gezien dat het onderscheid tussen de drie types interactoren precies ligt in de mate waarin er een direct verband bestaat tussen M/E en I (dit verband is symbolisch voorgesteld door de verbindingslijn tussen M/E en I in onderstaande figuur). Daar waar het verband tussen M/E en I heel direct is voor Alfa-attractoren, zijn de bij de informatie in Gamma-interactoren een zeer specifieke interpretatie nodig.
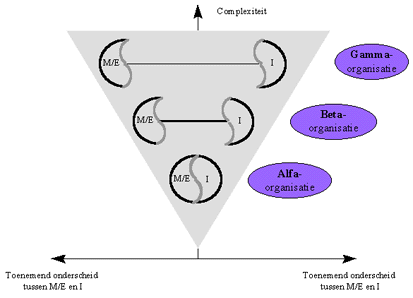
Figuur H10F10 : complementariteit tussen M/E en I als vorm van intregratie tussen M/E en I.
Algemeen kunnen we met betrekking tot de complementariteit van informatie en materie/energie nog 2 speciale kenmerken definiëren, namelijk zelfbevordering en circulariteit.
- Complementariteit en zelfbevordering
Deze complementariteit tussen M/E en I werkt ook "zelfbevorderend". Doordat de M/E en I immers perfect op elkaar afgestemd zijn, zullen ze er gedurende de ontwikkeling van het evolutiepatroon voor zorgen dat juist die M/E en I ontstaat en/of geselecteerd wordt, die complementariteit waarborgen. Varianten op deze beproefde complementaire combinaties hebben minder kans op succes, en zullen daarom door een proces van natuurlijke selectie geleidelijk aan verdwijnen. Alle evolutiepatronen die we rondom ons waarnemen, hebben hun bestaan te danken aan succesvolle attractoren met zelfbevorderende kenmerken. De bioloog Richard Dawkins heeft hiervoor de term "selfish genes" gelanceerd of "egoïstische genen". Hij kwam op dit concept uit vanuit het besef dat het enige waar de genetische informatie op uit is, is zichzelf in zijn voortbestaan te bevorderen. Bijna alle levende wezens die we kunnen waarnemen, zijn voorbeelden van soorten met een efficiënt egoïsme. De niet zelfbevorderende genen hebben immers minder kans op overleven, en zijn gedurende de evolutie uitgerangeerd. In complexe levende systemen, zoals een menselijk bedrijf, is de zelfbevordering heel expliciet aanwezig. Zelfbevordering kan daarom aanzien worden als een "verbeterde vorm" van de eerder neutrale "zelfreflectie". Immers, in de evolutie zal een niet-zelfbevorderend systeem het steeds moeten afleggen tegenover een zelfbevorderend systeem.
Nochtans komen in de evolutie ook succesvolle vormen van altruisme voor. Zo zijn er bijvoorbeeld veel dieren die in groep leven. Een mooi voorbeeld hiervan zijn de zebras. De reden waarom de zebras in groep leven en elkaar beschermen, is echter precies ook omdat dit voor hen de meest succesvolle strategie is op lange termijn. Door in groep te leven, is de kans groter dat ze snel predatoren kunnen waarnemen, en snel kunnen vluchten. Voor de predatoren (zoals leeuwen of luipaarden) is het bovendien moeilijker om een zebra uit een groep te selecteren en aan te vallen. Dit betekent dat altruïstisch gedrag voortkomt, precies omdat het ook het voortbestaan van de soort beter bevordert op lange termijn. Ook altruïstisch gedrag is dus in wezen een kenmerk van de egoïstische genen. Bovendien komt dit groepsgedrag van de zebra overeen met wat we op lagere niveaus in de natuur hebben gezien. Onder invloed van stresscondities (wat het vooruitzicht op een roofdier zeker is), zullen de egoïstische interactoren de stress trachten te minimaliseren. Als gevolg hiervan zullen ze veelal een collectief gedrag vertonen.- Complementareit en circulariteit
Een ander gevolg van de toenemde zelfreflectie tussen materie/energie enerzijds en informatie anderzijds is een toenemende "circulariteit".
Door de co-evolutie worden alle systemen en processen meer en meer aan elkaar aangepast. Dit betekent dat een variatie van de ene interactor, kan leiden tot een variatie van een andere interactor. Dit kan opnieuw gevolgen hebben voor de kenmerken van de eerste interactor, enz...
De adaptatie of aanpassing van de systemen wordt dus in de eerste plaats beïnvloed door de onmiddellijke omgeving en de gebeurtenissen in het onmiddellijke verleden. Het is te vergelijken met een spel tafeltennis, waar de volgende slag, voornamelijk bepaald wordt door de vorige.
Wel is het zo dat de spelregels bij tafeltennis in het verleden zijn vastgelegd, en dat het volledige spel zich binnen deze regels afspeelt. De evolutie is te vergelijken met een spel tafeltennis tussen M/E en I, waarbij de regels zelf geleidelijk aan evolueren. De spelers kunnen dus beslissen om met drie te gaan spelen, om de tafel te vergroten of het net te elimineren. Daarom is het zeer moeilijk te voorspellen hoe het spel zal aflopen.
Om die reden heb ik de notie van "gemiddelde evolutie" geïntroduceerd. Als we de spelers honderdduizend maal laten spelen, kunnen we misschien wel bepaalde terugkerende spelregels of gewoontes vaststellen. Deze spelregels zijn dan de "attractoren" van de gemiddelde evolutie.
Deze spiraal van steeds toenemende complexiteit, met een toenemende diversificatie van materie,energie en informatie, kan echter enkel onstaan ten koste van een toenemende verspreiding.Complementariteit tussen M/E en I is dus een essentieel aspect is van attractoren.
De meeste wetenschappelijke vakgebieden leggen echter ofwel de nadruk op het energetische of materiële aspect van de evolutiepatronen, ofwel juist op het informatie-aspect, veelal met nadruk op de tegenstelling tussen M/E en I.Filosofen richten zich traditioneel op het probleem van de scheiding tussen "lichaam" (M/E) en "geest" (I) op een zeer abstracte wijze, die ervoor zorgt dat het probleem minimimalizeerd wordt. Biologen zouden een sterke interesse moeten vertonen voor het principe van de complementariteit tussen M/E en I, omdat alle biologische organismen in grote mate afhangen van informatie voor de controle van hun energie- en materiestromen. Nochtans trachten meer "traditionele" biologen zich in de meeste gevallen van het probleem af te houden door een sterk reductionistische visie, waarbij de voornaamste inzet gericht wordt op het materie/energie-aspect van de biologische wezens, door bijvoorbeeld de genetische code van het DNA te ontrafelen. De recente bloei van de "bio-semiotiek" als relatief nieuwe tak van de biologie, geeft hier wat tegenwind. Fysici daarentegen maken een strikte scheiding tussen materie/energie enerzijds en informatie anderzijds door het gebruik van universele wetmatigheden, die betekenis krijgen door het aangeven van de juiste contextuele informatie (de zogenaamde begincondities). Nochtans zijn er in de fysica duidelijke voorbeelden van interactie tussen M/E en I. In quantumfysische experimenten kan bijvoorbeeld aangetoond worden dat het uitvoeren van een meting (dit is het verwerven van informatie over een M/E situatie), de situatie op zichzelf beïnvloedt. Meten is dus niet enkel weten : meten is ook beïnvloeden. Andere vakgebieden, zoals bijvoorbeeld de artificiële intelligentie of het artificieel leven, benadrukken dan weer heel sterk de informatiekant van de realiteit, en meer bepaald van het levende systeem.
Het is precies door het invoeren van het concept van de Alfa-, Beta- en Gamma-attractoren, met elk hun relatie tussen M/E en I, dat ik in dit boek getracht heb om de complementariteit tussen M/E en I meer op een voetstuk te plaatsen. Door het invoeren van het concept van de attractoren met complementaire M/E en I, is het mogelijk een meer consistente beschrijving te geven van de evolutie, over de diverse vakgebieden geen. Dit was geen doel op zichzelf, maar een middel dat zich gaandeweg heeft ontwikkeld.
Terug naar de index van dit hoofdstuk
5. De evolutie lijmt haar eigen scherven
De evolutie is gekenmerkt door een reeks opeenvolgende symmetriebrekingen, waarbij de symmetrie van het universum steeds verder gebroken wordt.
Deze algemene tendens van symmetriebreking wordt echter op verschillende plaatsen verstoord door het ontstaan van attractoren. Deze zorgen voor het onstaan van interactoren die wel een gemiddelde of individuele symmetrie vertonen, die ingaat tegen de algemene tendens.
Bovendien evolueren deze attractoren zodanig, dat de interactoren een toenemende reflectie vertonen
Figuur H10F11 : Invloed van divergerende trend (gevolg van verspreiding) en convergerende mechanismen (als gevolg van de fluxmaximaliserende principes) in de gemiddelde evolutie (nota bene : de rechten wijzen allerminst op een "lineair verband", ze tonen gewoon de trend aan).
In het proces van de co-evolutie ondergaan de attractoren en de interactoren van eenzelfde niveau en van verschillende niveaus een gezamelijke evolutie.
Doordat hierbij ruimte en tijd meer en meer geïntegreerd worden, reflecteert het heden meer en meer zowel de gebeurtenissen uit het verleden, als deze in de toekomst.
Bovendien is spontane zelforganisatie van attractoren gekenmerkt door een complementariteit van materie/energie en informatie, is ook hier een steeds toenemende reflectie waar te nemen tussen de materie/energie en de informatie .Het lijkt wel alsof in de evolutie continu "scherven" worden gemaakt door de opeenvolgende symmetriebrekingen en differentiaties. Wat we echter ook vaststellen is dat deze evolutionaire scherven ook regelmatig "gelijmd" worden door een tegengestelde beweging van integratie en reflectie.
Het is wellicht precies door deze twee tegengestelde doch complementaire bewegingen dat het universum gedurende de evolutie, ondanks de overmacht van "De Verspreiding", toch als samenhangend en harmonieus wordt ervaren.
Bovendien kan men zich terecht de vraag stellen hoever deze opeenvolging van symmetriebreking en reflectie kan doorgaan.
Men kan immers volgende redenering maken : als de evolutie interactoren voortbrengt die een steeds toenemende vorm van reflectie vertonen van het verleden en de toekomst in het heden, kan dit proces extreem ver doorgaan. Uiteindelijk ("ad infinitum") kunnen er evolutiepatronen onstaan die zelf de volledige evolutie kunnen repliceren, inclusief het verleden en de toekomst. In dit geval heeft het evolutieproces zichzelf voortgeplant. Het proces der processen, namelijk de evolutie, zou aanleiding geven tot een ultieme zelfreflectie, als gevolg van het ontstaan van een "Universele replicator". "De Big Bang baart een baby" . Dit zou betekenen dat de Big Bang op zich model staat voor een evolutionaire attractor (de "Oer-attractor" of in het Engels zou men kunnen zeggen "the Big Attractor"), van een hoger type dan de Alfa-, Beta- en Gamma-attractoren.Een volgende vraag die men zich dan kan stellen is : "De hoeveelste generatie is ons universum waarin wij ons bevinden ?". Wat we nu het begin van de evolutie noemen, is dan immers maar een herhaling van een bepaald proces. De herhaling van de evolutie zou dan geen virtueel gedachtenexperiment zijn, doch een realiteit. De attractoren in "de gemiddelde evolutie" zouden dan werkelijk kunnen bepaald worden.
Wat er ook van zij, beste lezer, op dit moment is dit een zuiver product van de verbeelding.
Verbeelding echter van een levend systeem, de mens, die toch al heel wat van het verleden en de toekomst in het heden kan beleven ...
In elk geval kunnen we met redelijke zekerheid stellen dat de gemiddelde evolutie "het midden houdt" tussen een aantal extreme toestanden, zoals :
- het streven naar chaos of streven naar orde
- het streven naar integratie of differentiatie
- innovatie of confirmatie
- instabiliteit of stabiliteit
- symmetriebreuk of coherentieWanneer de evolutiestroom zich een weg baant tussen deze zelfgeschapen en multidimensionele rivierwanden, kan de complexiteit van de interactoren en attractoren geleidelijk aan toenemen.
Hierbij lijkt er een convergentie op te treden naar een stromingsgebied waar er een evenwicht is tussen kanalisatie en vrij stroming van materie, energie en informatie. Een hoge complexiteit doet zich enkel voor op een grensgebied tussen orde en chaos, waar een balans gevonden wordt tussen innovatie en confirmatie van de informatiestroom.
Waar aanvankelijk De Verspreidende of divergerende trend in de evolutie oorspronkelijk zegeviert als alleenheerser, wint de evolutiestroom geleidelijk aan "zelfzekerheid". Deze "reductie van onzekerheid" kunnen we waarnemen onder de vorm van verschillende types informatie. Zijstromen van verschillende bronnen gaan meer en meer samenkomen en convergeren. Door deze groei van de coherente evolutiestroom , in kwantiteit en kwaliteit, wordt de flux van de stroom meer en meer gemaximaliseerd.
De gemiddelde evolutie-stroom mondt daardoor uit in een harmonie tussen divergentie en convergentie.