![]()
![]()
![]()
The formation of the gastrula in small holoblastic eggs results from the activity of a simple epithelium (blastula wall, one cell thick). Gastrulation is itself a process whereby the basic plan of the organism is established as a tube within tube, endomesoderm within an envelope of ectoderm. There is an initial flattening, and often a thickening, of the vegetal region of the blastula to form the vegetal plate, and then pushing of this plate as an invagination. The early embryos of many echinoderms, brachipods, enteropneusts, and Amphioxus, undergo form changes almost entirely by expansion and folding of a simple epithelium, much as sketched above (Fig.1).
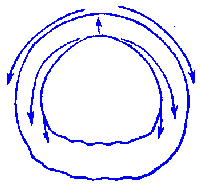
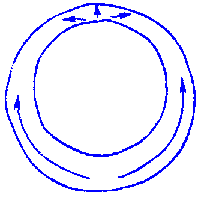
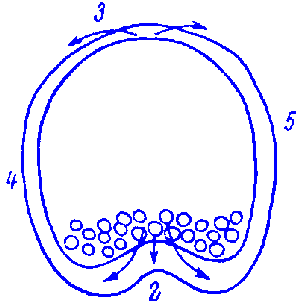
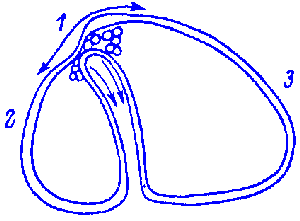
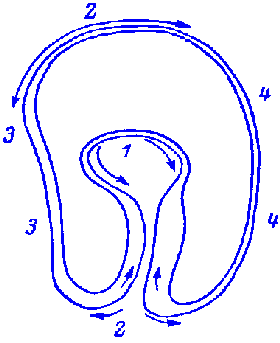
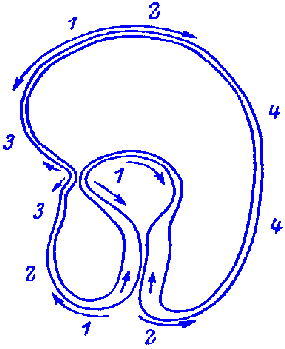
FIG.1. Stages of early embryogenesis of echinids
show development as interplay of physiological gradients (indicated by
arrows) and following morphogenetic movements (Sketched according to Child,
1936). Ciphers indicate relative levels of the gradients.
As Wilt (1987) described, the events resulting in specification of cell lineage are a necessary prelude to actual morphogenesis that forms a sea urchin embryo. The activities of the individual cells and their responses to cues that transcend domains of single cells drive the formation of the embryo. These morphogenetic activities in turn entrain further refinements in the definition of the pattern of differentiation. It is appropriate to inquire how determination in early stages causes morphogenetic movements and how these movements in turn lead to further pattern specification and differentiation.
Rather the picture here is of a very complex, highly correlated cellular machinery. According to Cummings (1994), this machinery must include and connect the correlated process of the genome, the mechanism for orchestration of cyto-skeletal elements or mechanism responsible for cellular shape, as well as the connections between these various elements. Epithelial sheets can remain as connected entitles while folding, invaginating, and deforming to produce a variety of three-dimensional structures. The linkage of cells into epithelia provides a means of converting mechano-chemical cell activity into particular kinds of larger scale mechanical motion and reordering, with important consequences for form and inductive signaling.
The different genetic and epigenetic events must apparently be reconciled but how this happens in detail is not known. What seems to be required is a place-dependent dynamic regulatory loop that ranges from the gene, to the gene products, to the cell layers, and back from the structures of higher scale to control the expression of the same gene or a different gene (Edelman, 1988).
![]()
![]()
![]()