
|
|
Photosystem 0: PS0 |
Since they use the same ATPsynthase, combined MTS/PS0 systems can be imagined, but they are not further discussed, although such systems could be relevant for the evolution of PS0.
Many aspects of chloroplast physiology can be related to PS0, in particular State transitions.
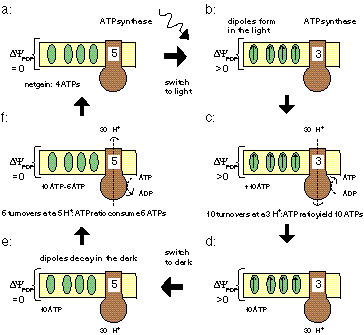
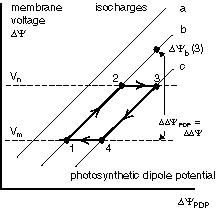
PS0 makes use of light-induced dipoles that decay in the dark. Such dipoles are
commonly found in socalled inactive PSII reaction centers,
where quinone QA cannot reduce quinone QB.
Illumination of such inactive centers is known to increase the membrane potential [Chyl89,
90].
Many other types of metastable light-induced dipoles are found during photosynthesis, and can result
in phosphorescence, luminescence, delayed fluorescense and thermoluminescence.
Fluctuating light
Objects carried along by convection currents will move in and out of the photic zone, they will be light cycled.
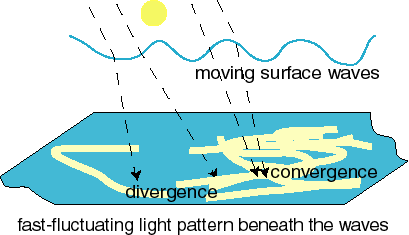
In other environments other types of light cycling occur. Due to light focussing effects by surface waves,
light fluctuations occur just under the surface of natural waters
[Sche57, Dera67,
70,
Snyd70,
Wein76].
They have a typical cycle time of 100 ms, although slower and
faster fluctuations occur as well, yielding a range of 0.01 - 1 s
[Queg86,
Wals83,
88],
which compares favorably with the range for thermal cycling times, > 10 s.
|
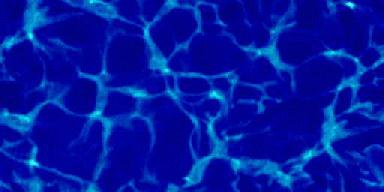
These light fluctuations are the cause of the complex light
pattern seen on the bottom of swimming pools.
Such patterns are often favorite backgrounds in movies or on TV, especially for
advertisements or titles: Algae Free Australia ad |
|
|
The evolution of PS0
There have been many studies on the evolution of photosynthesis
[Gran57, Gaff62,
Olso70,
87a,
87b,
Bord75, Gest80,
Sche83, VanG87, Pier89, Awra92, Blan92, Boxe92,
Hart92, Mauz92, Lark93]. These
studies typically consider charge transfer across the membrane instead of the formation
of light-induced dipoles within the membrane.
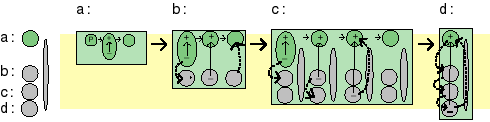
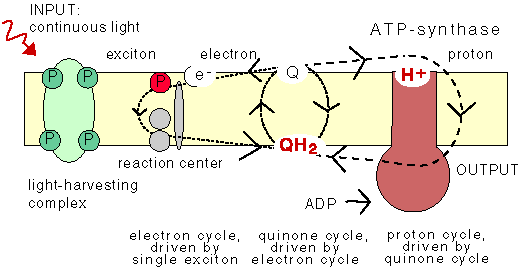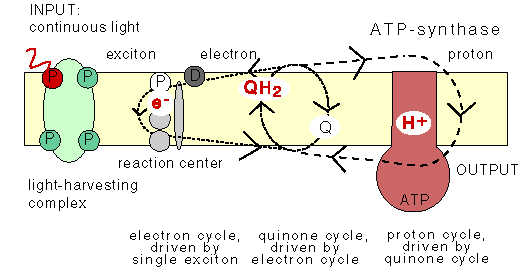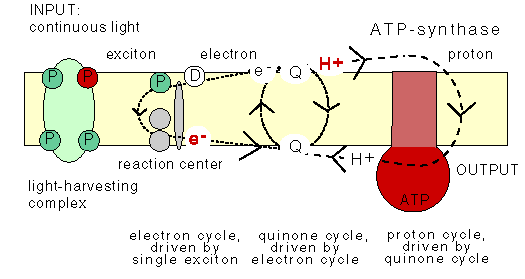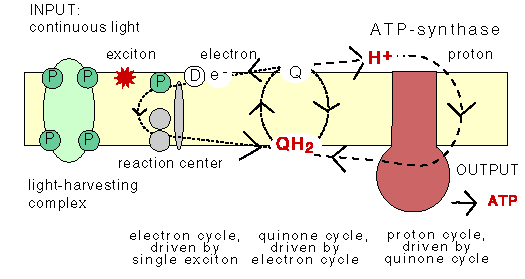
In PS0 the evolution of photosynthetic reaction center is conceptually very simple: temporarily charge carriers are added one after the other. This results in an array of stepping stones for excited electrons across the membrane.
The addition of quinones
By adding quinones that upon reduction can detach from the reaction center, pick up protons from
the medium, diffuse across the membrane, reject the protons and electrons, and can diffuse back through
the membrane, a system has been obtained that can perform in chemiosmosis in the standard way.
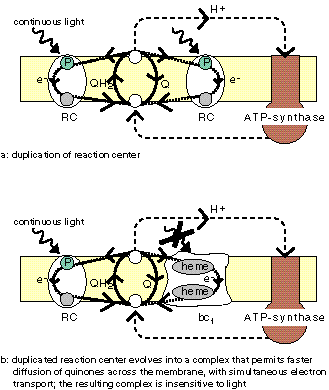
The Q-cycle
The animation does not show the Q-cycle, in which
the reduced quinone, after having diffused across
the membrane, returns one electron to an oxidized pigment,
and the second electron to a quinone at the other
side of the membrane using the bc1 protein which contains
several cytochromes as stepping stones. The Q-cycle can only be active
at low ĵH+ across the membrane.
|
It has been proposed that bc1 descends from a reaction center [VanG87], but no homology with reaction center proteins has been reported. |
Submissions to scientific journals are rejected all of the time. The 1995 paper on Photosystem 0 is rather long, as I perceived a need to discuss the many components of PS0 in detail. The referee did not see a 'fundamental problem' with the paper, and even called the idea 'interesting' but he called it 'overkill', suggesting a shorter version.
My motivation for the indeed unusually long paper was the large explanatory power of PS0. Many quantitative values are not too large, nor too small, but 'just right'; moreover, many puzzling phenomena can, admittedly to a variable degree, be linked to Photosystem 0.
In 2003 I submitted a shortened version to another journal. This paper was rejected as well, but in this case the referee misunderstood the idea.
Discussion
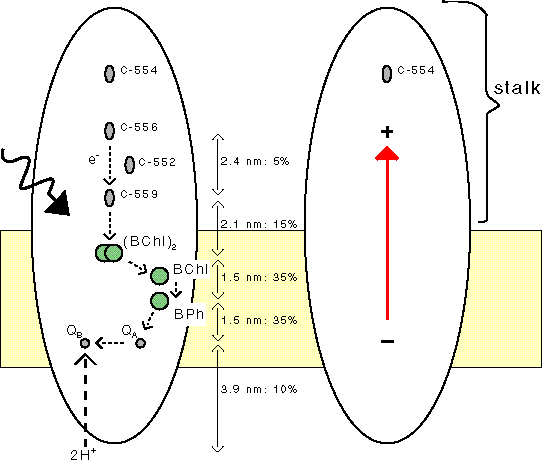
Many photosynthetic bacteria contain a stalk that
extends to far out the membrane. In Photosystem 0
this stalk has a function, as it constitutes a
large dipole. In regular photosynthesis the stalk has no function at all.
The chloroplast shows many phenomena that reminds
one of PS0: etiolation, stacking and swelling, induction, oscillations, and State transitions.
Terrestrial environments in which thermosynthesis and photosystem 0 could occur are discussed on: Terrestrial niches for thermosynthesizers.
Copyright © 1999-2005 Anthonie W.J. Muller