Ancha Srinivasan a, C. Johansen b, *, N.P. Saxena b
a Regional Science Institute, 4-13, Kita 24 Nishi 2, Kita-ku,
Sapporo, 001, Japan
b International Crops Research Institute for the Semi-Arid
Tropics, Patancheru 502 324, A.P., India
Received 26 April 1997; revised 14 October 1997; accepted 17 October 1997
Abstract
Most chickpea cultivars grown in regions where temperatures fall below 10oC continue to flower but fail to set pods. Research was conducted to characterize cold stress in a subtropical northern Indian location, Hisar and to examine genetic variation in pod and seed set at low temperatures. Lines of various maturity groups were used so that phenological timing of stress differed with line. Flowering was therefore synchronized with a cold spell to evaluate tolerance. A substantial variation was found both in field and controlled environments. Two early maturing advanced breeding lines, ICCVs (ICCV = ICRISAT Chickpea Variety) 88502 and 88503, showed good pod set in cold spells, but seed growth in them was limited. Evaluation of the parents indicated possible contribution of a germplasm from the former Soviet Union, K 1189 (ICRISAT chickpea germplasm collection No ICC 8923), to cold tolerance. Based on experiments in controlled environments, it is suggested that (a) pod set can occur at night temperatures of 0-5oC if daytime temperatures exceed 20oC (b) seed development requires a higher threshold temperature than pod set and (c) 15/5 and 15/0oC (day/night) thermal regimes are useful in preliminary screening of germplasm lines for cold tolerance. ã 1998 E1sevier Science B.V. All rights reserved.
Keywords: Chickpea (Cicer arietinum L.); Cold
tolerance; Pod set; Northern India; Genetic variation; Germplasm screening;
Flowering
1. Introduction
Prevalence of low temperatures (0-10oC minimum) during early flowering leading to excessive floral abortion is a major cause of low pod and seed set in chickpea (Cicer arietinum L.) in subtropical South Asia (Saxena, 1980). Other factors such as high humidity (Sengupta and Roy, 1979), cloudy weather (Chandrasekharan and Parthasarathy, 1963) and wet soil (Saxena and Johansen, 1990) are also implicated in poor pod set. Therefore, the improvement of cold tolerance, defined here as the ability to set pods and seeds at low temperatures, is of considerable significance. Besides having ability to set pods and seeds during cold spells, cold-tolerant lines are likely to have other advantages such as reduction of excessive vegetative growth leading to less lodging, reduced pest and disease incidence and greater harvest index (Saxena et al., 1988; Saxena and Johansen, 1990).
Three important requirements in the development of cold-tolerant lines
are (a) characterization of stress (b) identification of genetic variation
and (c) availability of simple screening methods. The objectives of this
study were to characterize stress experienced by field-grown chickpea and
relate it to the extent of floral abortion, assess genetic variation for
cold tolerance both in field and controlled environments and define a thermal
regime to facilitate rapid screening of chickpea germplasm and breeding
lines.
2.1. Field experiments
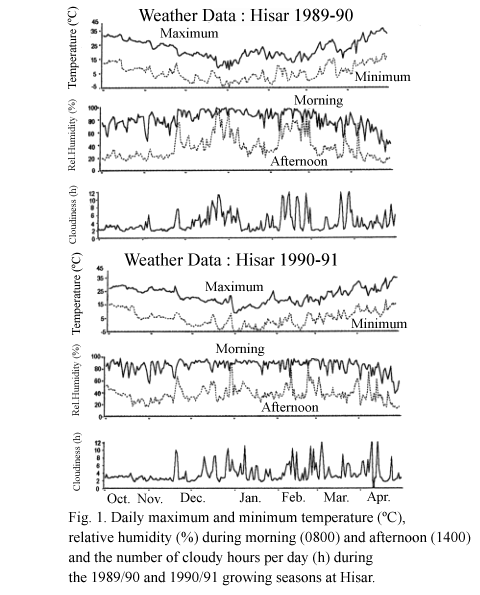
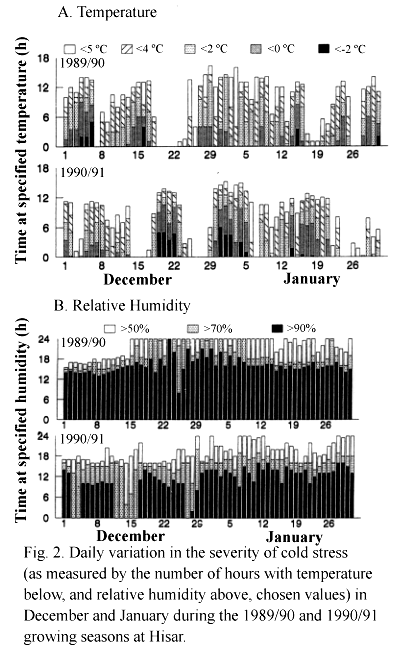
The experiments were conducted at ICRISAT Cooperative Center, Hisar (29o10'N, 75o46'E, 215 m above sea level) m northern India during 1989-90 and 1990/91, on a well-drained Entisol that was left fallow in the preceding rainy season. The climate of the site is subtropical monsoonal with a mean annual rainfall of 447 mm. The maximum and minimum screen air temperatures, relative humidity at 0800 and 1400 and number of cloudy hours per day during the period of experimentation are shown in Fig. 1. Diurnal changes in temperature and RH at canopy level were monitored from November to March using mini-thermohygrographs (Cole-Parmer Instrument, Chicago, IL). The severity of cold stress was measured by the number of hours with temperature below, or RH above, chosen values.
In 1989/90, two traditional cultivars (Pant G 114 and Gaurav) and 14 lines derived from crosses between a kabuli germplasm (K 1189) from the former Soviet Union and Indian desi germplasm were sown on 27 October, 20 November and 5 December 1989 (Table 1). In 1990/91, the same set of 16 cultivars/lines was sown besides Annigeri, Chafa, G 130, K 1189 and Pant G 115. Flowering in all cultivars/lines was synchronized with the cold spell by grouping them according to phenology and planting each set on an appropriate date and by artificially extending daylength to 18 h to hasten flowering. The experiments were conducted in a split plot design, with planting dates as main plots and cultivars/lines as subplots and replicated 3 times. Subplots in 1989/90 were 24 m2 in area while in 1990/91 they consisted of three 6.0 m long rows with an inter-row spacing of 30 cm. Plant population was in the range of 25-28 plants m-2. In both seasons, the entire area received 10 t farmyard manure (FYM)+ 10 kg N + 20 kg P (as diammonium phosphate) ha-1 prior to the first planting. A pre-sowing irrigation was given to ensure uniform and rapid emergence. Rhizobial inoculation was not done, as responses are not found in this area. Nodulation occurred on all cultivars/lines. Seedlings were thinned to 10 cm between plants at 15-20 days after sowing. The experimental area was hand-weeded and the crop protected from pests and diseases, as necessary.
2.1.1. Data collection and sampling
Dates of anthesis (first flower opening) and first pod set in eight randomly chosen plants per subplot were recorded. In 1989/90, flowers, which had opened during December and in the first and second fortnights of January and the pods set, were counted separately in five plants per subplot. In 1990/91, however, such counts were grouped at 10-day intervals in 10 selected cultivars/lines. The tolerance of a cultivar/line was determined on the basis of the proportion of pods formed out of the number of flowers opened during the cold spell. In 1990/91, cold tolerance was scored on a l-5 scale (1, > 75%; 2, 50-75%; 3, 25-50%; 4, 5-25%; 5, <5% pod set) during the cold spell after observing > 90% floral abortion in a susceptible cultivar (Annigeri).
2.2 Experiments in controlled environments
The experiments were conducted at ICRISAT, Patancheru, using walk-in growth rooms (Models CG811 and CG1011, Controlled Environment Canada) and growth cabinets (Percival Manufacturing, IO). Growth cabinets were used to impose 0oC during night. Growth rooms had 72% input wattage of 1500-mA cool white fluorescent and 28% input wattage of Sylvania 50W-277V incandescent lighting. The average irradiance (400-700 nm) at the top of the canopy was 320-340 m mol m-2 s-1 (LI-COR IA 188B Photometer /Radiometer, LAMBDA Inst. Lincoln, NE). The light and dark periods were 12 h each with an abrupt change. The air temperatures varied less than ± 1oC around the set temperature, as sensed with a shielded thermocouple. The soil temperatures were not controlled but measurements at the center of the pots (8-10 cm deep) showed that they equilibrated at prevailing air temperatures within about 3 h. The RH was 65 ± 5% during the day and 90 ± 5% at night, as measured with mini-thermohygrographs. Airflow rates over the top of the canopy, measured using an OMEGA HH-F10 air speed indicator, varied between 0.37 and 0.45 m s-1.
2.2.1. Cultural practices
Plants were grown in 2 l volume, white-painted pots containing a mixture of Vertisol soil, sand and vermiculite (2:2:1 by volume). Six seeds were sown in each pot and inoculated with Rhizobium (IC 59). Seedlings were thinned to three after expansion of the first multi-pinnate leaf. Plants were alternately supplied with 35 ppm N solution (as ammonium nitrate) and tap water, the amount and time of application varying depending on thermal regime, while ensuring that all cultivars/lines within a regime were treated equally. Pots were randomized on alternate weeks to minimize any positional effects.
2.2.2. Choice of thermal regimes
Based on the reports of Summerfield et al. (1981) and Roberts et al. (1985), 25/15oC (day/night) was chosen to represent the control regime. The other temperature combinations were selected to represent a range typical of those experienced by chickpea during early flowering in northern India. Plants were grown in the control regime until 4-5 days before anthesis (judged on the basis of size of the first formed bud) and allocated to different thermal regimes. After a 30-day exposure, plants were transferred back to the control regime.
Initially, only six cultivars/lines (Annigeri, ICCVs 88502 and 88503 in the early maturity group and Pant G 114, ICCVs 885l0 and 88514 in the late maturity group) were evaluated because of space constraints in growth rooms. In a preliminary experiment, all six cultivars/lines (three pots each) were grown in both rooms with a single regime (25/15oC day/night) to evaluate cultural practices and to quantify differential room effects. In another preliminary experiment, responses of cultivars/lines ex-posed to pre-anthesis long day treatment (i.e., 18/6 h day/night for 20 days after seedling emergence and grown at 12/12 h thereafter) were compared with those of plants exposed to 12/12 h photoperiod continuously, to examine whether long days altered pod set responses significantly.
The main experiment was conducted in two cycles over time, with both cycles repeated to test reproducibility of results. In cycle 1, five pots i.e., 15 plants) each of six cultivars/lines were subjected to 25/15, 25/5, 20/5 and 20/0oC regimes. In cycle 2, plants were exposed to 25/15, 15/5, 25/0 and 15/0oC regimes. All 21 cultivars/lines included in the field trial in 1990/91 were later tested for pod and seed set in 15/5 and 15/0oC regimes. Plants transferred to cool regimes tended to develop branches at the lower nodes. In order to reduce diversion of assimilates to vegetative organs, new axillary growth that appeared was removed in all plants.
2.2.3. Data collection and statistical analysis
Several buds were tagged and observed daily to determine the duration of pod development. The number of pods was counted and the proportion of pod set determined. The plants were harvested when most pods had dried. Analyses of variance were done according to standard procedures (Steel and Torrie, 1980).
3. Results
3.1. Characterization of cold stress in the field
Three key elements in characterization of an abiotic stress are intensity and duration of stress, rate of stress development and phenological timing of the stress. Because standard meteorological data (Fig. 1) did not correctly reflect changes in the microclimate, temperature and RH measured at the canopy level were used to characterize cold stress. In both seasons, cold stress was severe m December and January. There were more cold days in 1989/90 than in 1990/91, but the days on which minimum temperature fell below -2oC was more in the latter (Table 2). Diurnal fluctuations of temperature and RH in December and January revealed that both seasons were characterized by several cold spells, interspersed with warm periods (Fig. 2). For example, in 1990/91, cold stress was severe during the third week of December and in the first week of January, with temperature being below 0oC for about 10-12 h in a day. Cold spells started later in 1989/90 than in 1990/91. Ground frost was not noticed in 1989/90 but it was observed on 4 successive days from 17 December in 1990/91. The difference between day and night temperatures and the number of hours with high relative humidity (>70%) were higher in 1989/90 than in 1990/91. The change in temperature over time (rate of stress development) did not follow any consistent trend. Cold stress was thus variable in occurrence and intensity and hence unpredictable under field conditions.
The severity of stress experienced by cultivars/lines varied with their maturity group and date of sowing. In 1989/90, the early maturing lines sown in October passed through the cold spell during early flowering while the late maturing ones did so during late vegetative growth. The November and December-sown cultivars/lines passed through the cold spell largely during vegetative growth. In view of this, only the results pertaining to the October sown crop are discussed here. In 1990/91, however, all cultivars/lines passed through the cold spell during early flowering due to adjustments in sowing date and imposition of long day treatment.
3.2. Genetic variation in pod and seed set under field conditions
In 1989/90, the seed yields of cultivars/lines sown in November were, in general, greater than yields of those sown in October or December (Table l) as most of the cultivars/lines sown in October produced a heavy canopy and lodged, while those sown in December had poor vegetative growth and reduced seed filling due to a sudden rise in temperatures later in the season.
The analysis of flower, pod and seed production patterns in the October-sown crop during December l989 and January 1990 (Table 3) revealed several points. Of the cultivars/lines that commenced flowering in December and passed through the cold spell, pod set was > 5o% in ICCVs 88501, 88502 and 88503 only. The number of pods with developing seeds was, however, very small. The pods were either empty or contained small shriveled seeds. Seed growth was slightly greater in ICCV 88503 than in others. Late-maturing cultivars/lines commenced podding in relatively warm periods. A few cultivars/lines such as Pant G 114 and ICCV 88510 set pods in the last week of January probably due to unusually warm daytime temperatures, but they failed to set pods during the cold spell in the first week of February. Thus, due to fluctuating temperatures and differences in time to anthesis, cold tolerance could not be precisely evaluated in the late-maturing cultivars/lines.
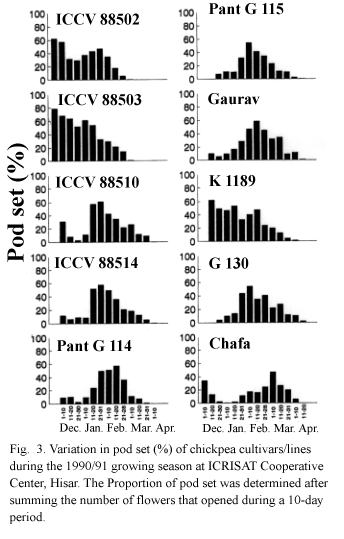
In 1990/91, all cultivars/lines passed through the cold spell during early flowering. The analysis of flower, pod and seed set patterns in 10 selected lines of contrasting phenology and sensitivity to cold stress (Fig. 3) showed that ICCVs 88502 and 88503 had a very good ability to set pods in cold spells, as earlier observed (N.P. Saxena, ICRISAT unpublished data). Late maturing lines such as ICCVs 88510 and 88514, which were earlier selected as possible cold tolerant lines but with some uncertainty as to whether they had merely escaped cold stress (N.P. Saxena, ICRISAT, unpublished results), failed to set pods in the cold spell. They were indeed as sensitive to cold as the control cultivars. However their rates of flower production (i.e., number of flowers opened per day), pod set (i.e., number of pods set per day) and pod growth in the later part of January and early February were greater than in other lines (data not shown). This could be a major reason for their greater seed yields recorded in the previous years (ICRISAT, unpublished data, 1988). Of all six parents, only K 1l89 set a few pods in cold spells thereby indicating its possible contribution to cold tolerance.
The observations from field experiments in both seasons showed that no line bore pods continuously at all nodes in cold spells. This indicates that low temperature under field conditions interfered with normal podding of all cultivars/lines tested.
3.3. Experiments in controlled environments
All chickpea cultivars/lines in 25/15oC regime grew well and set pods normally. Analysis of variance for seed yield and dry matter showed no significant (P > 0.05) room effects. Replication of thermal regimes was therefore considered unnecessary. Instead, the entire experiment was repeated to test reproducibility of results. Cultivars/lines exposed to pre-anthesis long-day treatment commenced flowering simultaneously at 30-35 days after emergence, while those in the 12/12 h (day/night) treatment started flowering at different times. Annigeri and ICCVs 88502 and 88503 flowered around 30-35 days, while Pant G 114, ICCVs 885l0 and 88514 flowered around 95, 80 and 90 days respectively. Pod set responses were not, however, significantly affected by photoperiod treatments. It was therefore considered appropriate to expose plants to long days in the later experiments as the treatment would save time and enable simultaneous comparison of cultivars/lines.
3.3.1. Effects of temperatures and genetic variation in pod set
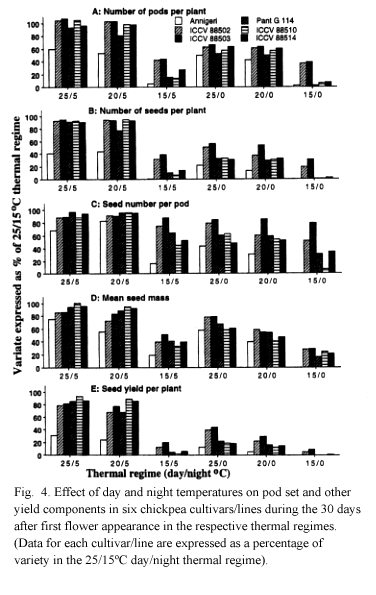
Data in different thermal regimes were standardized by expressing them as a percentage of the value in the control (25/15oC) regime. In this way, the relative sensitivity of cultivars/lines could be directly compared. In the control regime, the late maturing cultivars/lines grew taller, accumulated more dry matter and set a few more pods than the early maturing lines, as expected (Table 4). A declining trend in pod set was observed in response to decreasing temperatures in all cultivars/lines (Fig. 4a). However, contrary to expectations, even sensitive cultivars/lines such as Pant G 114 set pods normally in regimes with low night temperature (25/5 and 20/5oC). A drastic reduction in pod set at these regimes occurred in Annigeri, however, thereby indicating its greater sensitivity to low night temperatures. A moderate pod set was observed in all cultivars/lines even at 25/0 and 20/0oC.
Distinct genetic variation in pod set was, however, noticed at 15/5 and 15/0oC regimes. At these regimes, pod set was moderate (approximately 40% of the control regime) in ICCVs 88502 and 88503 only. The poor pod-setting ability of the late maturing lines, ICCVs 88510 and 88514, at these regimes indicated their sensitivity to low temperatures and this result corroborates the earlier observations from field experiments.
The conclusions regarding pod set are more or less similarly applicable to other yield components (Fig. 4b-e). In all cultivars/lines seed yield (Fig. 4e) and its components (Fig. 4b-d) were less in regimes with 0oC than in those with 5oC night temperature. Seed development responses at 15/5 and 15/0oC regimes, where genetic variation in pod set was distinct, indicated a drastic reduction in seed growth even in tolerant lines, ICCVs 88502 and 88503. In ICCV 88502, both seed number per pod (Fig. 4c) and mean seed mass (Fig. 4d) were reduced. In ICCV 88503, however, the reduction in mean seed mass was greater than in seed number per pod, thereby indicating a negative influence of low temperature on dry matter accumulation in seeds.
The ranking of 21 cultivars/lines in low temperature regimes was similar to that in the field experiment (Table 5) as the rank correlation coefficients both in 15/5 (r = 0.87***) and 15/0oC (r = 0.84***) regimes were significant at 0.001 level of probability.
4. Discussion
The failure of chickpea flowers to set pods in cool climates constitutes wastage of sink capacity. Although such loss is considered an adaptive mechanism that stimulates vegetative growth and provides additional nodes/sites for subtending more flowers and pods (Saxena, 1984), development of cold tolerance has several associated advantages (Saxena et al., 1988; van Rheenen et al., 1990). As tolerant lines can set pods early in the season, vegetative growth is likely to remain moderate, thereby limiting chances for lodging in the later stages. Incidence of foliar diseases may be reduced and the crops can escape from Helicoverpa damage. The lines may also escape terminal drought and heat stress, as they are likely to complete the life cycle early. All these advantages lead to a high harvest index and yield stability and make such lines ideal for adaptation to environments similar to that of Hisar and for introduction into new cropping systems (e.g., chickpea-sugarcane in north-western India) and new regions such as in Western Australia.
Low night temperature was earlier implicated as a major cause for floral abortion in field-grown chick-pea (Saxena, 1980) as satisfactory pod set could be obtained by soil heating cables and polythene cover which increased ambient temperature around the crop (Saxena, 1984). Based on such studies, pod set was reported to occur only when minimum night temperature exceeds 8oC (ICRISAT, 1988; Saxena and Johansen, 1990). Our studies in controlled environments, however, suggest that pod set response arises from the net effect of both day and night temperatures rather than from low night temperature alone, because moderate pod set was noticed in sensitive lines even when night temperature was 0oC (as long as the daytime temperature was around 20oC or more). The response was not mediated by diurnal temperature however, as wide differences were found at 15/5 and 20/0oC. That even cold sensitive varieties were able to set pods at 0 and 5oC suggests that a higher daytime temperature of around 20oC may partly compensate the negative effects of low night temperature. These results are in agreement with those of van Schaik and Probst (1958) and Lawn and Hume (1985), who reported that warm daytime temperatures compensated for cold nights in soybean. Lyons (1973) also cited several examples wherein it was shown that chilling injury could be avoided in many tissues if they are returned to a warm temperature before degenerative changes occur.
The enhanced floral abscission at 15/5 and 15/0oC regimes is particularly relevant as such temperatures are not uncommon under field conditions. Day and night temperatures quite often drop below 15 and 0oC respectively for at least a few hours especially in December and January in many chick-pea-growing regions of northern India and these could be deleterious to pod and seed set. As ranking among the cultivars/lines at 15/5 and 15/0oC in controlled environments reasonably matches with that in the field during cold spell (Table 5), the thermal regimes may be very useful in preliminary screening of germplasm. A greater reduction in pod set at l5/5 and 15/0 than at 20/5 and 20/0oC regimes respectively may also be interpreted to mean that lower daytime temperatures induce floral abortion. Grantz (1989) had suggested that cool temperatures persisting into daytime contribute to photoinhibition.
While it is reasonable to conclude based on these studies that low pod set was due to the combined effects of low day and night temperatures, separation of the effects of low temperature and high RH was not possible here, as growth rooms did not have a provision for humidity control. RH values were indeed very high in December and January under field conditions especially in the 1989/90 season (Fig. 2b). High RH was earlier suspected to be a major cause for poor pod and seed set (Sengupta and Roy, 1979; Sindhu et al., 1981) as the success in chickpea cross pollination was higher when RH was in the range of 35-53%. High RH (> 50%) is known to prevent dehiscence of anther sacs (Sen and Mukhejee, 1961).
Notwithstanding the uncertainty regarding the role of temperature and RH in pod set, the evidence from both field and controlled environment studies clearly shows that there is a substantial genetic variation for cold tolerance. While the number of cultivars/lines tested here is obviously small, the variability in response is reasonably large. The cultivars/lines may be selected for use in different agroecological zones, depending on their level of tolerance. For example, while ICCVs 88502 and 88503 may be useful in regions with moderately severe winters as in northern India, others such as ICCVs 88506 and 88509 may be relevant in mild winters of central India. The cultivars/lines tested may be grouped as those with ability to set pods in (a) Severe cold spell: ICCV 88503 (b) Mild cold spell: ICCVs 88502, 88501, K l 189, ICCVs 88506, 88509, Gaurav - in that order (c) Moderately warm periods: Pant G I 14, ICCVs 88514, 885l0, 88511, 88505, 88512, 88504, Pant G 115, G 130, ICCVs 88515, 88508, 88513 and (d) Very warm to moderately hot periods: Annigeri and Chafa.
While substantial genetic variation was seen in pod set, seed growth in all cultivars/lines including ICCVs 88502 and 88503 was limited during cold spells under field conditions and at low temperatures in growth rooms. These results suggest that the critical temperature required for seed growth was higher than for pod set and thus point to the need to screen germplasm for better seed development in cold conditions. Seed growth in all cultivars/lines was indeed much better in pods set in the warm periods of late February than those set in the cold spells of December or January under field conditions. This observation corroborates the later findings in controlled conditions, where pods set in the warm regime seemed to be preferentially stronger sinks than those set in the cool regimes, irrespective of the location of flower on the plant (Srinivasan, A., unpublished results).
Extreme caution is necessary however, in extrapolating responses in controlled environments to the field. The thermal regimes in the field fluctuate and are never similar to those in controlled environments. The interaction with other factors such as soil moisture status is probably more important under field conditions (Saxena and Johansen, 1990). Further, the root temperatures experienced in controlled environments may be different from those experienced by their counterparts in the field at similar air temperatures. Duke et al. (1979) reported that normal pod set occurred in soybean when root temperature was raised. Plants in controlled environments here were transferred abruptly into the cool regimes without any period of acclimation. Refinements such as de-signing a reasonably dynamic temperature regime, simulating that in the field, might be expected to improve the predictability of field responses.
The implication of the results for development of chickpeas for regions subject to chilling stress is clear. It is emphasized that breeding strategies must seek to develop lines with a greater degree of adapt-ability to cool climates. Towards this end, the physiological and biochemical basis for cold tolerance must be clarified further. If a specific reproductive process limiting pod and seed set is identified as more sensitive to low temperature than the others are, selection for that trait may improve the overall efficiency of breeding programs. The role of factors other than low temperature must also be looked into. For example, the effects of low light intensity associated with cloudy weather under field conditions are unclear. Studies to determine the importance of such factors are described in subsequent papers.
We thank Dr. L. Krishnamurthy, Dr. H.S. Talwar, Mrs. Y. G. Devi, Mr. Raj Singh and other Crop Physiology staff for their assistance. We also thank Dr. S.C. Sethi, former Research Coordinator and Manager at ICRISAT Cooperative Center, Hisar, for facilitating the field trials.
Chandrasekharan, S.N., Parthasarathy, N,, 1963. Botany of useful plants-Indian gram. Madras Agric. J. 16, 5-1 1.
Duke, S.H., Schrader L.E., Henson, C.A., Servaites, J.C., Vogelzang, R.D., Pendleton, J.W., 1979. Low root temperature effects on soybean nitrogen metabolism and photosynthesis. Plant Physiol. 63, 956-962.
Grantz, D.A., 1989. Effect of cool temperatures on photosynthesis and stomatal conductance in field-grown sugarcane in Hawaii. Field Crops Res. 22, 143-155. ICRISAT, 1988.
International Crops Research Institute for the Semi-Arid Tropics Annual Report 1987. Patancheru. A.P., India, pp. 136-137.
Lawn, R.J., Hume, D.J., 1985. Response of tropical and temperate soybean genotypes to temperature during early reproductive growth. Crop Sci. 25, 137-142.
Lyons, J.M., 1973. Chilling injury in plants. Ann. Rev. Plant Physiol. 24, 445-466.
Roberts, E,H., Hadley, P., Summerfield, R.J., 1985, Effects of temperature and photoperiod on flowering in chickpeas (Cicer arietinum L.). Ann. Bot. 55, 881-892.
Saxena, N.P., 1980. Pod setting in relation to temperature at Hissar. Int. Chickpea Newslett. 2, 11.
Saxena, N.P., 1984. Chickpea. In: Goldsworthy, P.R., Fisher, N.M. (Eds.), The Physiology of Tropical Field Crops. Wiley, New York, pp. 419-452.
Saxena, N.P., Johansen, C., 199O. Realized yield potential in chickpea and physiological considerations for further genetic improvement. In: Sinha S.K., Sane, P.V., Bhargava, S.C., Agarwal, P.K. (Eds.), Proc. Int. Congress Plant Physiol., Vol, I, 15-20 February 1988, New Delhi, India. Indian Soc. Plant Physiol. and Biochem. New Delhi, India pp. 279-288.
Saxena N.P.. Johansen, C., Sethi, S.C., Talwar, H.S., Krishnamurthy, L., 1988. Improving harvest index in chickpea through incorporation of cold tolerance. Int. Chickpea Newslett. 19, l7-l9.
Sen, S., Mukhenjee, D., 1961. Preliminary studies on defective seed setting in gram. Sci. Cult. 27 (4), 185-188.
Sengupta K., Roy, P.N., 1979. Factors affecting success of cross pollination in chickpea. Indian J. Agric. Sci. 49, 489-491.
Sindhu. J.S., Lal. S.B., Singh, R.P., 198l. Studies on the factors determining crossing success in chickpea (Cicer arietinum L.). Pulse Crops Newslett.. 1, 21-22.
Steel, R.G.D., Torrie, J.H. 1980. Principles and Procedures of Statistics. McGraw-Hill, New York, 481 pp.
Summerfield, R.J., Minchin, F.R., Roberts, E.H., Hadley, P., l981. Adaptation to contrasting aerial environments in chickpea (Cicer arietinum L.). Tropical Agric. 58 (2) 97-114.
van Rheenen, H.A., Saxena N.P, Singh, K.B., Sethi, S.C., Acosta-Gallegos, J.A., 1990. Breeding chickpea for resistance to abiotic stresses: what are the problems and how can we solve them? In: Chickpea in the Nineties: Proc. Second Int. Workshop on Chickpea Improvement, 4-8 December 1989. ICRISAT Center, India. International Crops Research Institute for the Semi-Arid Tropics, Patancheru, pp. 239-243.
van Schaik, P.H., Probst, A.H., 1958. Effect of some environmental factors on flower and reproductive efficiency in soybeans. Agron. J. 50, 192-197.
{kind=link}
{kind=link}
{kind=link}
{kind=link}