![]()
![]()
![]()
Classical embryological analysis has lead to the concept of a morphogenetic field. It can be defined operationally as the domain within which regulation can occur in response to surgical manipulation (French, Bryant and Bryant, 1976). In several organisms it has been shown that up to certain stage the whole embryo can regulate and it therefore constitutes a single primary field. But later surgical interventions have more localized effects, restricted to developmentally autonomous parts of the embryo. These units French, Bryant and Bryant (1976) called secondary fields.
French, Bryant and Bryant (1976) carried out a set of key experiments and correlated their findings on very diverse epimorphic regenerating systems (cockroach legs, Drosophila imaginal discs and newt limbs) with their elegant polar coordinate model. The model accounts formally and in a simple and unified way for the kind of developmental regulation seen in the secondary fields of both invertebrates and vertebrates.
This is as contrasted with "morphollactic" regeneration which take place in the absence of cell division but rather by modeling of existing tissue, and in the present terms, assigning a new value of morphogen. A common example of morphollactic regeneration occurs in the hydra. The situation of the epimorphic regeneration of "imaginal discs" of insects (from which many of adult appendages such as legs, antennae, wings, sex and mouth parts arise) , which take place after surgical intervention of various kinds, is summarized below.
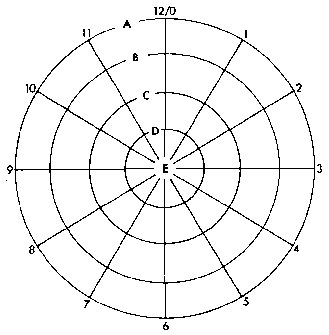
Polar coordinate model. Each cell is assumed to have information
with respect to its position on a radius (A through E) and
its position around the circle (0 through 12). According
French, Bryant and Bryant (1976).
The following two rules for the behavior of cells in epimorphic fields are sufficient for regulations.
Application of the model to the results obtained by culturing imaginal disk fragments from Drosophila. According French, Bryant and Bryant (1976).
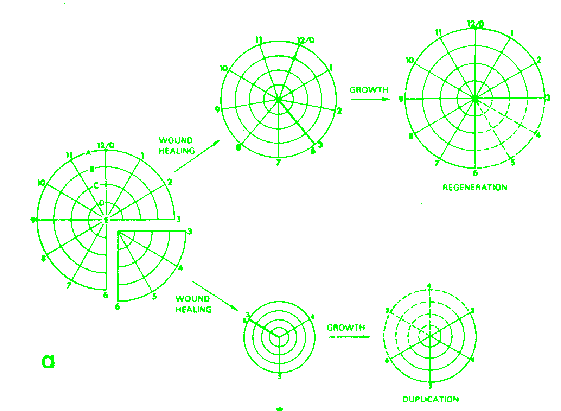 Results
of culturing 90 or 270 degrees sectors of the wing disk.
Results
of culturing 90 or 270 degrees sectors of the wing disk. 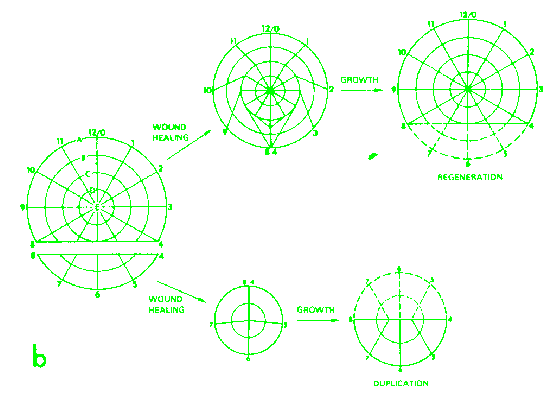 Results
obtaining following bisection of the wing disc.
Results
obtaining following bisection of the wing disc. 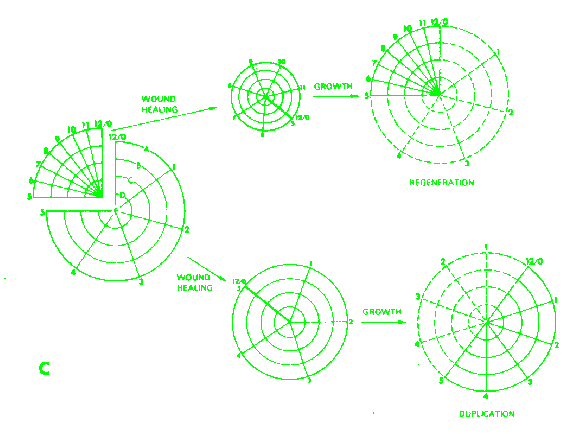 Results
of culturing different fragments in the leg disk.
Results
of culturing different fragments in the leg disk. 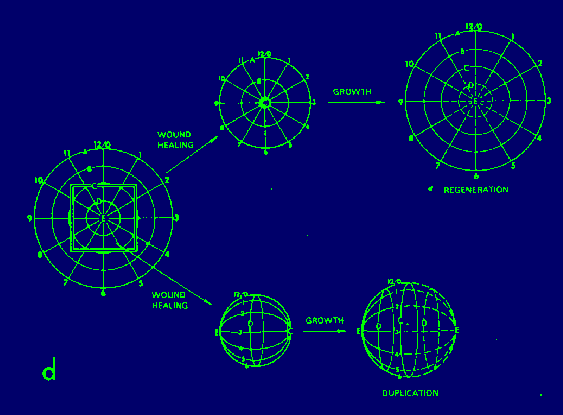 Results
obtained by culturing central and peripheral fragments of the bilaterally
symmetrical genital disk.
Results
obtained by culturing central and peripheral fragments of the bilaterally
symmetrical genital disk.
![]()
![]()
![]()