
|
|
Introduction |
The TS theory may be testable with
little effort and expense in today's organisms, but conversely TS may also have occurred such a long ago that no traces of it have remained. In the latter case it would remain of interest because of the theoretical solution to the problem of the origin of life.
The origin of life, the early evolution of biological energy conversion, and the early evolution of photosynthesis still lack general accepted models. How could such models be developed? Which assumptions can be made, which working methods can be used? We cannot go back in time, and the course of evolution cannot directly be studied.
The 'tree of life' is commonly used in evolution models. The development from simple to complex, the 'real tree' must remain a closed book, but
several projections of it have been considered. There are many 'trees of life': the the real tree, the extant tree, the fossil tree, and the proposed functional tree.
Contemporary, 'extant' organisms have been classified according to complexity in what we shall call the 'extant tree'. Organisms considered as the most complex (mammals, ourselves) are placed at the top and the most simple, unicellular extant organisms at the bottom.
Computer programs permit the establishment of the degree of kinship between extant organisms from their genomes, yielding an 'extant molecular data tree'. Enhanced horizontal gene transfer between the genomes of early organisms (such as during endosymbiosis, widespread in protists [Marg81]) may have intertwined the early branches of the real tree so much that its root cannot be traced any more.
Fossils of extinct organisms yield upon arrangement the 'extinct fossil tree'. Not all organisms have left fossils, and the information therein is also limited.
It is uncertain whether, and if so, under what conditions, nucleic acids remain preserved in fossils: no 'extinct molecular data tree' has been described, although there has been much recent progress in analysing old DNA.
In order to follow the evolution of physiological processes from
the very beginning, the origin of life, one runs into the problem, clearly
recognised by Darwin, that a first organism that arose today would
probably be quickly devoured by contempary organisms. In that case the extant tree cannot
cover the origin of life. The origin may also similarly fall outside
the area covered by backwards extrapolation of the
extant tree, the extant molecular data tree, and the extinct fossil tree.
The extant molecular data tree shows that the last common ancestor of all living organism was most closely related to the Archaebacteria living today in boiling hot springs
[Woes87], an environment conducive to convection.
Studying the extant physiology tree, for instance of photosynthesis, may therefore not resolve the physiology of the origin of photosynthesis. Can we look elsewhere?
The issue of finding the solution to the origin of life may very well be a question of imagination, not one of experimenting on something that lies in front of us. The first concepts will have to be from the inanimate world. It may be a question of design. An 'engineering approach' is proposed; we try to find a "blueprint for life" [DeDu91].
We therefore look outside the box of the life sciences, and consider man-made physical devices, or 'machines' or 'engines.' Some life scientists may deride implicating machines in the origin of life as 'speculation' that may be never be able to proof anything. But this method can yield new viewpoints, new ideas, which suggest new experiments, even on extant organisms. New ideas are essential for scientific progress: as clearly recognised by Darwin, before a scientist starts to observe or to experiment, he should have an idea to test his results against.
Machines may not only explain the origin of photosynthesis, but their mechanisms may still occur in present organisms, having only therefore not been observed, as the eye can only see what's in the mind, which in turn is mostly familiar with the phenomena that have been already been observed and explained. Such an approach is no idle speculation, where new theoretical concepts are needed. In Blake's words [Blak90]: "What is now proved was once, only imagine'd."
The engineering approach has many advantages. The origin of life occurred by definition in an inanimate world. Engineers tend to take a close look at energy and thermodynamics, are used to thinking in different time scales and in terms of function. Man-made machines show a diversity and complexity comparable to that of organisms. The engineer has no qualms of being speculative, he does not apologize for being imaginative. Where the properties of parts are understood, he feels no hesitation to combine them in at first imaginary devices.
The origin of life model given in this document is certainly feasible from an engineering point of view. The testing of a proposed biological mechanism supposedly similar to an device constructed by engineers may be difficult but, again, feasible. If the functions that one wants to perform or to observe are well-defined, the appropriate experiment is often easy.
Heat engines
Every document on thermodynamics discusses heat engines. See for instance:
Encyclopaedia Britannica on Thermodynamics
The steam engine is the best known example of
a heat
engine. In the boiler, steam is formed that pushes a piston, which does
the
sought-for external work. In the condenser cooling water takes up heat
from the
expanded steam, which condenses and is next pumped back into the
boiler. This internal water therefore circulates within the engine, and is thermally
cycled.
According to the First Law of Thermodynamics heat is not a conserved entity, but can be converted into work. The sum of heat and work,
the energy, is however conserved. The external work obtained from the engine therefore equals the difference between the heat absorbed from the fire by the boiler room and the heat rejected to the cooling water in the condenser. 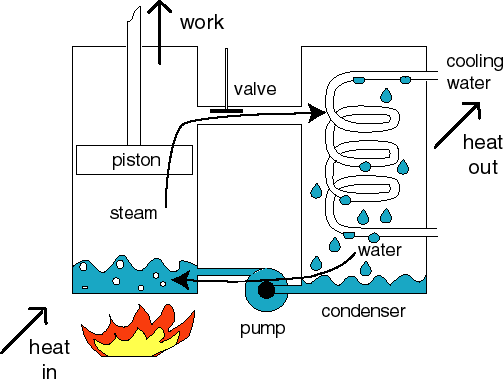
The change in volume and pressure
of the water within the steam engine: 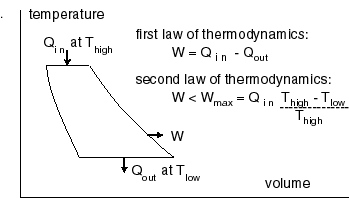
Many types of heat engines are known. They may make use of other
fluids than water, or may even not use a fluid at all, such as in thermoelectricity. All heat engines take up
heat at a high, and reject it at a low temperature.
Directly after the elucidation of the laws of thermodynamics in the last century Helmholtz, Thomson, Boltzmann and Frick considered the possibility that the muscle functions as a heat engine [Flor72, Brod75].The numbers do however not add up. From the known efficiency of biological energy conversion in, for instance, muscle and photosynthesis the minimal high temperature of the involved heat engine can be estimated. Such estimates yield implausible temperatures above 100°C. Moreover, significant heat conduction from the hot to the cold reservoir cannot be avoided in a muscular heat engine.
As a consequence, many contemporary studies state explicitly that biological heat engines are impossible. The following citation from a biology textbook [Vill85] is typical:
Many of the machines used in industry are heat engines. An
engine driven by steam produced by the burning of coal in a boiler is a familiar
example of a heat engine. However, heat is not a useful way of transferring or
storing energy in biological systems. Under conditions of constant pressure heat
can do work only when it can flow from a region of higher temperature to a
region of lower temperature. Living organisms are basically isothermal
(equal-temperature) systems; there is no significant temperature gradient-that
is, difference in temperature-among the various parts of the cell or the various
cells in a tissue. Cells cannot act as heat engines, for they have no means of
permitting heat to flow from a warmer to a cooler body.
This statement overlooks the many cases where organisms are present in a thermal gradient: thermoclines in natural waters, many interfaces, the leaf illuminated by the sun, convecting hot springs. Moreover, the statement may not apply to the origin of life at all. Other authors are more careful [Darn86, p 20]:
For heat to do work, it must flow from a region of higher temperature
to one of lower temperature. Differences in temperature often exist between the
internal and exteral environments of cells; however, cells generally cannot
harness these heat differentials to do work.
Another author [Voge88, p 308]:
As far as we know, nature has never evolved a heat engine. The "why nots" of evolution may be truly unknowables, but one might venture a guess here. Organisms carry out their activities in a temperature range extending from the freezing point of water up to where their proteins become chemically or structurally unstable. On top of any other inefficiencies, a biological heat engine would have to suffer a thermodynamic "tax" of around 90% as a consequence of equation [the Carnot ratio]. And a primitive version using a lower temperature difference would have even greater inefficiency."
The counterargument of a low efficiency is not valid. It was probably derived by only taking into account the establishment of a thermal gradient by the 'burning' of food, whereas nature also shows many thermal gradients which could be used directly. The low Carnot efficiency does not make TS implausible. The first steam machines had a very low efficiency as well. What must have been important for an early biological energy source was not efficieny but simplicity, ease of construction. Note that PTS makes use of the mentioned "structural instability" of proteins.
Nature's columnist Deadalus has proposed biological heat
engines in 1997 [Jone97]. He
pointed out that in several environments "Carnot life" might be possible, but
did not propose a specific mechanism.
He mentioned the requirement for a
"weird metabolism".
Most heat engines are built by man but the convection
cell, also a heat engine, can emerge spontaneously, it can
self-organize.
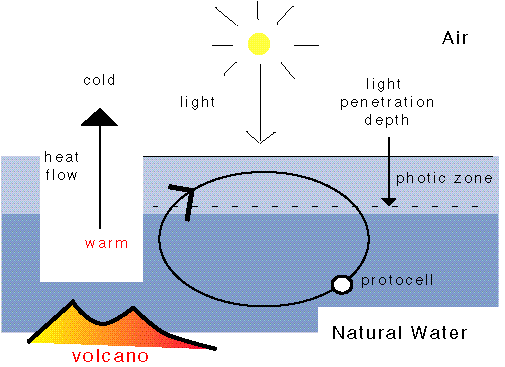
What is convection?
Smoke rises since its temperature is above that of the surrounding air; its lower density makes it float in the surrounding air. After cooling smoke descends again.
In a
closed space, say a room with a heat source like a stove, and
in which air is cooled near the windows, air similarlyt circulates: from stove to ceiling to windows to floor, back to the stove. This circulation during heat transfer in a fluid
- gas or liquid - is called convection.
In 1900 Benard did the first detailed experimental study of the spatial pattern of fluid flow during convection; important contributions were later made by Rayleigh [Kosc93 p 4]
|
Literature on convection: Introductions: Encyclopaedia Britannica on Convection [Hake78, Chap91] Specialized monographs [ Chan61, Kosc93] |
Convection
may seem an almost trivial and unimportant phenomenon. But it is quite profound. Most of the order in the world around us is the result of
convection. Convection occurs on many length and time scales. It transports heat
from the interior of the Sun, where at a temperature of millions of degrees
nuclear fusion occurs, to the Sun's surface. On Earth, irradiation by the Sun
causes large scale convection in both the atmosphere and the ocean, driving all
weather phenomena and all sea currents. In the Earth's mantle, convection drives
plate tectonics.
These convection processes in turn drive other processes: weather drives the hydrological cycle of
ocean > water
evaporation > rain > river > ocean
which again drives several
geological processes. Similarly, plate tectonics results in earthquakes and volcanism, which also drive other additional processes. Ore formation is an example of ordering that results from multiple processes, all driven by
convection.
Convection is self-organizing and can do external
work. A windmill is driven by the weather caused by convection. While doing work there is always some free energy loss, or
dissipation, due to friction; any system that does external work is a dissipative structure. Any process requires an energy source, and the question
arises what the energy source of the origin of life could be - what is the
relevant dissipative structure?
'In the conclusion of his first paper Benard referred to the interest that the hexagonal cells had found among the "naturalists". This interest has continued until today. However the obvious similarity between Benard cells and living cells has yet to prove a fruitful connection. In view of the complexity of living cells and the difficulties which had to be overcome in order to explain even the "simple" Benard cells, it is not surprising that a real connection between both phenomena has not yet been made.'
|
|
|
In TS the precise spatial patterns set up during convection are
unimportant, as use is made of the thermal cycling of the fluid
during its circulation.
Davies (1998) similarly mentions convection in the context of
the origin of life as an example of self-organization, but sees no possible
link. First he declares (probably referring to the Lava lamp):[Davi98 p 6] |
The first
organisms are in the TS theory under the control of the convection current.
External thermal cycling can control many partial systems and yields within a
composite system a simple method for synchronization of the functions of the parts.
TS gives a solution for the control problem mentioned by Davis, giving a path for a stepwise transition of
external to internal control.
Conclusion
Convection has been related to the origin of life by many authors but only in very general terms. The thermosynthesis theory describes a detailed coupling between convection and the origin of life.
The next page gives details of this coupling, showing how the thermal cycling during convection could have driven primordial bioenergetics by a heat engine-like mechanism: Protein-associated Thermosynthesis.