
|
|
The origin of life |
Recent reports mention the synthesis during extraterrestrial conditions of amino acids on ice crystals under UV light [Bern02, Muno02]. They have also been detected in meteorites [Maut95], and have also been found in Miller's classical experiments [ Mill53, 55]. The presence of amino acids on the primitive Earth is therefore plausible.
More generally many small biomolecules have been found in outer space recently as well [Ehre00, Char00].
Step 2: Large biomolecule synthesis
Bernal has proposed the formation of polypeptides from amino acids during the drying
of tidal pools at low tide and at mild temperatures (~50°C) [
Bern67].
Proteinoids
Polypeptides
Proteinoid microspheres
The emergence of the genetic system
Fox has demonstrated condensations of amino acids during dry conditions
just above the boiling water of water (~120°C)
[
Fox59,
72,
80,
81,
88,
91;
Laha78]. Fox called the
products proteinoids. Proteinoids can have molecular weight as large as 6000. They should
Recently a new variation of polypeptide synthesis has been reported by Matsuno. The peptide bonds is formed
when water is split off from the -COOH and -NH2 groups present in two different aminoacids or peptides.
Given for instance two amino acids, the peptide bond reaction can occur twice, leading to the
formation of a diketopiperazine, which has a ring structure.
At first sight diketopiperazine seems a showstopper for peptide elongation. Matsuno has
however observed that
an amino acid can react with several diketopiperazines: the rings can be broken up, and
combined to form a long elongated polypeptide.
The reaction would occur during hydrothermal vent conditions [
Futa??].Step 3: Protocell formation
vesicles
Vesicles have been observed in primordial material [Maut95, Dwor01]. Such compartmentalization can help in selection processes, and may offer some protection of larger biomolecules against for instance hydrolysis.
Step 4: Convection
It follows from the Second law of thermodynamics
that the self-organization of life is only possible by invoking a dissipative
structure [
Hake78, preface], which as life did not existed yet,
must be found in the inanimate world .
Few inanimate dissipative structures are known.
An interesting one is the maser or microwave laser (for a discussion
of the laser as a dissipative structure, see [
Hage78]). In interstellar gas clouds
masers form spontaneously [
Elit95], and one may wonder whether such a maser could somehow
drive the origin of life. Other dissipative structures can arise from solutions of
chemicals that are not in equilibrium: the dissipative structures manifest
themselves as chemical oscillations. Such oscillations have been related
to oscillations in Ca2+ content within organisms.
The most common
self-organizing dissipative structure is however the convection cell.
Interestingly, Paecht-Horowitz has invoked convection
in the origin of life [
Paec84].
Convection is possible in a fluid, that is in a gas as well as a liquid.
Several studies ([
Sage94]) have considered primordial reactions in a convecting gas.
Aerosol and cloud formation would be the relevant processes.
For the purpose of this study, water is taken as
the convecting fluid. A
volcanic hot spring does function as the convection cell. The intensity of convection increases with
the temperature difference across the cell, and by having an ice cover of the spring a large temperature
difference is assured. In the literature there is some discussion on whether life started
at a high or at a low temperature. Evidence supporting both temperatures has been given. TS shows
that is no necessary contradiction, as a convection cell contains both high and low temperatures.
The surface ice means that snowball earth, an early Earth covered with ice,
is suited for the cradle of the origin of life.
Woese has already been proposed that the last common ancestor of all living
organisms lived in volcanic hot springs [
Woes87].
Step 5: Condensation during thermal cycling
The properties of the proposed pF1 have already been
mentioned. pF1 can perform PTS, i.e.,
it can condense several substrates resulting in phosphorylation or peptide
bond formation while it is being thermally cycled. The mechanism involves
the establishment of a local dehydrated environment within the enzyme, which
helps the product to form. The product can only be released by a thermal unfolding
of the enzyme. The mechanism thus is a thermal variation of the
binding change mechanism for F1 as described by Boyer and Penefsky.
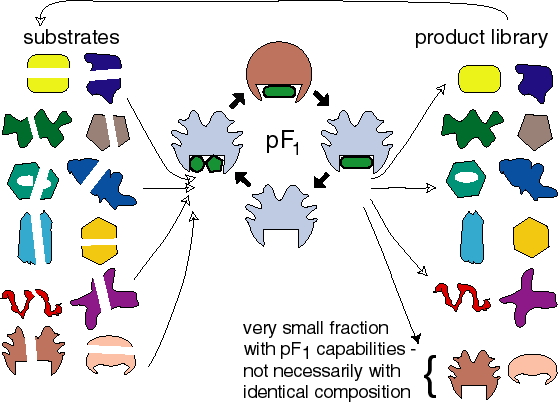
The library of products synthesized by pF1 contains itself some
molecules with pF1 capabilities.
The plausibility of the TS model for the origin of life depends crucially
on the probability of the chance that a molecule in the library is a pF1 synthesis.
Orgel has claimed that such a process is highly unlikely [
Orge87]. The use
of randomly constituted protein libraries is now standard practice in the
commercial search for enzymes. By adding thermal cycling the chance of pF1 formation
could be experimentally determined. The chance is expected to increase with the salt concentration, as
the decrease of water activity associated with the latter must decrease the
free energy for a condensation reaction.
PTS capability may selects microspheres, as membranes are stabilized by phosphorylation
of the membrane components [
Davi58,
West87].
At the end of step 5 thus a very inefficiently propagating pF1 has been obtained
that has a metabolism, condensing substrates, that uses thermal cycling.
Step 6: Information formation
Generalising an idea of Dyson on the origin of RNA [
Dyso82,
85], it is proposed that the
first information carrying molecules did not give a selective advantage to its parent,
and that they were like parasites. They are present in that large part of the library of
pF1 products that does not have pF1 capabilities.
Some pF1 is assumed to have been able to duplicate or replicate these information
carrying molecules, which we give the name of pRNA.
Note the similarity with the PCR method for nucleic acid
duplication, which also makes use of thermal cycling.
How pRNA was replaced by RNA cannot be modelled at this moment. The origin of the
required ribose remains a major problem [
Shap86,
88].
Alternatives for RNA have been found such as peptide nucleic acids, which even
can hybridize with DNA [
Niel91]. The author is not aware of alternatives that have
all the capabilities of present day RNA, including its ribozyme activity.
So an assumption is made in step 6, the emergence of RNA. PTS is the energy source for its replication.
Step 7: Information procession
see preprint (2005):
"Thermosynthesis as energy source of the RNA World: a new model for the origin of life"
RNA perform three functions:
ribosomal RNA (rRNA) constitutes the major part of the ribosome, the place
where proteins are synthesized, transfer RNA (tRNA)
activates amino acids, and messenger RNA (mRNA), which contains the information for
the peptide composition.
In the TS theory the ribosome enhanced the fidelity of pF1 propagation.
In a gradual process the precision increased, until a system making use of the genetic code had
appeared [Hart75,
78,
Eige71,
77,
81,
Woes65,
77,
80a,
80b,
Woes87,
Dyso85].
Many reactions are assumed to have involved pF1, other reactions may have involved
ribozyme-like reactions.
By defining a simple function for the protein product, the plausibility of its synthesis is enhanced: the synthesis of a complex enzyme is not that probable.
Step 8: Information storage
Upon the acquisition of reverse transcriptase it has become possible to synthesize DNA.
Appropriate thermozymes are obtained derived from present thermozymes for DNA transcription.
The genetic apparatus completed
After the emergence of the genetic system the rest of evolution can follow its Darwinian path.
Gene duplication, enzyme diversification and enzyme specialization make the evolution of life simple.
Step 9: Conversion to isothermy
It would give an organims a selective advantage to have its enzymes being able to work
during at a constant temperature instead of having a thermal cycling requirement. During evolution thermozymes are therefore expected to
have lost this requirement. The loss may have been effected by making use of protein phosphorylation,
which can be considered as a method to mimic thermal cycling of a protein, and, for membranes,
by making use of Ca2+, which can increase the thermotropic phase transition of a membrane.
Some proteins may however have kept the thermal cycling requirement. Thermal cycling still
is a requirement for many physiological processes: germination, vcell division, flowering, budding.
Many organisms still are subject to thermal cycling, for instance our daily body temperature
varies cyclicly, in women a monthly cycle exists; many organisms perform daily vertical migrations
in natural waters, which because of a thermocline must often result in thermal cycling.
Discussion
The model for the origin of life is of interest for several reasons.
(1) It is often claimed that no comprehensive origin of life model is possible. TS shows that this is not true. TS is however not the only theory on the origin of life that claims to be comprehensive, there are others, see [
Laha01].
(2) Second, many organisms show a thermal cycling requirement for cell division or germination. In TS this requirement is interpreted as a possible relic of PTS.
(3) The origin of life must support a simple metabolism.
Dyson has proposed that biogenesis comprised two steps, a first biogenesis and a second biogenesis [
Dyso82,85].
The first biogenesis would involve a primitive metabolism, and is
identified with PTS. The emergence of the genetic machinery code
constituted the second biogenesis; this occurs during the gradual
increase in accuracy of pF1 synthesis under the
influence of information carrying molecules. Other models
for the origin of life postulate large networks of enzymes (Kaufman, Lahav)[
Kauf93,
Laha01]; the many required enzymes, and the large genome, make the proposed emergence of the genetic machinery however implausible.
(4) In the model proteins come first, and nucleic acids come later.
Proteins take care of the required energy,
information transfer capability is acquired later.
It is often assumed that nucleic acids
must have come present before proteins during the origin of life.
But energy is required for information transfer -
the computer has to be switched on before it can copy a file.
Therefore, the chicken must have come before the egg.
(5) Since thermal cycling occurs on the surface of almost all celestial
objects, and will also occur in convection currents
underneath surface ice when this covers these objects, the origin of
life could have happened almost everywhere in the Solar System.
The next page gives the details of a second mechanism for thermosynthesis, a mechanism that makes use of biomembrane: Membrane-associated Thermosynthesis.
Copyright © 1999-2004 Anthonie W.J. Muller