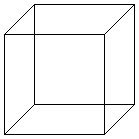
 Есть такая известная оптическая иллюзия - куб Неккера. Он представляет собой
рисунок из линий, который мозг интерпретирует как трёхмерный куб. Но существуют
две возможные ориентации этого мысленного куба, и обе они в равной степени
совместимы с двумерным образом на бумаге. Вначале мы обычно видим одну из этих
ориентаций, но если мы смотрим несколько секунд, то куб "перещёлкивается" в
мозгу, и мы видим другую очевидную ориентацию. Спустя ещё несколько секунд
мысленный образ щёлкает обратно, и эти перещёлкивания продолжаются, пока мы
смотрим на картинку. Суть в том, что ни одно из двух восприятий куба не является
правильным или "истинным". Они оба правильны. Точно так же то видение жизни,
которое я защищаю, и которое обозначаю термином "расширенный фенотип", возможно
не "правильнее" ортодоксальных воззрений. Просто это другой взгляд, и я
предполагаю, что он позволяет достичь более глубокого понимания предмета; по
крайней мере - в некоторых отношениях. Но я сомневаюсь, что возможны какие-то
эксперименты, которые могли бы доказать моё предположение.
Есть такая известная оптическая иллюзия - куб Неккера. Он представляет собой
рисунок из линий, который мозг интерпретирует как трёхмерный куб. Но существуют
две возможные ориентации этого мысленного куба, и обе они в равной степени
совместимы с двумерным образом на бумаге. Вначале мы обычно видим одну из этих
ориентаций, но если мы смотрим несколько секунд, то куб "перещёлкивается" в
мозгу, и мы видим другую очевидную ориентацию. Спустя ещё несколько секунд
мысленный образ щёлкает обратно, и эти перещёлкивания продолжаются, пока мы
смотрим на картинку. Суть в том, что ни одно из двух восприятий куба не является
правильным или "истинным". Они оба правильны. Точно так же то видение жизни,
которое я защищаю, и которое обозначаю термином "расширенный фенотип", возможно
не "правильнее" ортодоксальных воззрений. Просто это другой взгляд, и я
предполагаю, что он позволяет достичь более глубокого понимания предмета; по
крайней мере - в некоторых отношениях. Но я сомневаюсь, что возможны какие-то
эксперименты, которые могли бы доказать моё предположение.
Все явления, которые я рассмотрю - коэволюцию, гонки вооружений, манипуляции хозяевами со стороны паразитов, манипуляции живыми существами неодушевлённым миром, "экономические" стратегии животных по минимизации затрат и максимизации выгод - достаточно знакомы, и уже являются предметом интенсивного изучения. Зачем же тогда читатель должен тратить своё время и усилия на продолжение чтения? Заманчиво позаимствовать обаятельно бесхитростный призыв Стивена Гулда в начале его значительного тома (1977a) и просто сказать: "пожалуйста, прочтите эту книгу", и вы узнаете, почему стоит тратить на неё усилия. К сожалению, у меня нет оснований для аналогичной самоуверенности. Я, будучи обычным биологом, изучающим поведение животных, могу лишь сказать, что воззрение, обозначенное как "расширенный фенотип" заставило меня увидеть животных и их поведение по-иному, и я думаю, благодаря этому я понимаю его теперь лучше. Сам по себе "расширенный фенотип" не может считаться проверяемой гипотезой, но он меняет способ оценки животных и растений, благодаря чему он может навести нас на такие проверяемые гипотезы, о которых мы в противном случае никогда не мечтали бы.
Открытие Лоренца (1937) о том, что поведенческий паттерн может быть уподоблен анатомическому органу, не было открытием в обычном смысле слова. В его подтверждение нельзя было привести никаких экспериментальных данных. Это был просто новый способ интерпретации фактов, который ныне стал уже банальностью, и до сих пор доминирует в современной этологии (Tinbergen 1963). Сегодня он кажется нам настолько очевидным, что нам трудно представить, что когда-то требовалось его "открытие". Точно так же знаменитая глава Д'Арси Томпсона (1917) "По теории трансформаций..." получила широкое признание как работа большой важности, хотя она не выдвигала и проверяла гипотез. В известном смысле очевидна строгая истинность того факта, что очертание любого животного можно преобразовать в очертания родственного животного посредством математического преобразования, хотя не очевидно, что преобразование будет простым. Практически выполняя это преобразование на множестве конкретных примеров, Д'Арси Томпсон навлёк на себя реакцию типа "ну и что?" всякого, достаточно скрупулёзно настаивающего на том, что наука продвигается только опровержением конкретных гипотез. Если мы прочтём главу Д'Арси Томпсона, а затем спросим себя, - что же мы теперь знаем из того, что мы не знали прежде, то разумным ответом будет: немного. Но наше воображение будет взбудоражено. Мы возвращаемся и смотрим на животных новым взглядом; мы уже думаем о теоретических проблемах, о новых путях эмбриологии, филогенеза и их взаимосвязей. Я конечно, не столь самонадеян, чтобы сравнить свою скромную работу с шедевром крупного биолога. Я привожу этот пример лишь для того, чтобы демонстрировать, насколько может быть стоящей для прочтения теоретическая книга, даже если в ней не выдвигаются проверяемые гипотезы, но вместо этого предлагается изменить способ видения рассматриваемого предмета.
Другой крупный биолог однажды рекомендовал для лучшего понимания фактической картины рассмотреть также и возможные варианты: "Ни один практический биолог, изучающий половое размножение, не станет вести работу по подробному исследованию жизни организмов, имеющими три или более полов; но тем не менее, что же он ещё должен делать, если хочет понять, почему в реальности полов всегда два?" (Fisher 1930a). Вильямс (1975), Мейнард Смит (1978a) и другие - показали нам, что одна из обычнейших, универсальнейших особенностей жизни на Земле - само наличие полов, нельзя принимать некритично. На деле - их существование решительно удивляет, если его противопоставить предполагаемой возможности бесполого размножения. Вообразить бесполое размножение как гипотетическую возможность нетрудно, ибо мы знаем, что оно действительно имеет место у некоторых животных и растений. Но существуют ли другие случаи, в которых наше воображение не получает такой подсказки? Имеются ли подобные важные факты о жизни, которые нам трудно заметить просто потому, что у нас не хватает воображения представить альтернативные варианты вроде трёх полов Фишера, но которые могли бы иметь место в каком-то возможном мире? Я постараюсь показать, что да.
Игра с вымышленным миром с целью лучшего понимания реального называется техникой "мысленного эксперимента". Мысленные эксперименты очень охотно используются философами. Например, в сборнике статей о "Философии мышления" под редакцией Гловера 1976, некоторые авторы воображают хирургические операции, в которых мозг одного человека пересаживается в тело другого, а они в ходе мысленного эксперимента выясняют значение "личностной идентичности". Бывает, что мысленные эксперименты философов чисто мнимы и безумно невероятны, но это не существенно с точки зрения цели их проведения. Но бывает, что они в большей или меньшей степени учитывают факты реального мира, например факты экспериментов по рассечению мозга.
Рассмотрим другой мысленный эксперимент, на сей раз из области эволюционной биологии. Однажды в студенчестве я должен был написать теоретический очерк о "происхождении хордовых" и о других вопросах древнего филогенеза. Один из моих наставников справедливо старался поколебать мою веру в ценность существующих построений, предлагая предположить, что что-нибудь могло в принципе, развиваться во что-нибудь другое. Даже насекомые могли развиться в млекопитающих, если бы должная последовательность давлений отбора происходила в должном порядке. В то время, как и большинство зоологов, я отклонил идею как очевидную ерунду; я и сейчас конечно же не верю, что нужной последовательностью давлений отбора было бы достаточно. И мой наставник - тоже. Но то, насколько принцип важен, почти неопровержимо показывает простой мысленный эксперимент. Нам нужно только доказать, что существует непрерывный ряд маленьких шагов, ведущих от насекомого, скажем - самца жука-оленя, до млекопитающего, скажем - оленя. Я имею в виду, что начиная с жука, мы могли бы составить такую последовательность гипотетических животных, близких к предыдущему члену последовательности как к родному брату, которая достигла бы высшей точки в самце королевского оленя.
Доказательство будет нетрудным при условии, что мы согласимся с тем (а с этим согласны все мы), что и жук и олень имеют общего предка, хотя и очень далёкого. Пусть даже не существует другой последовательности шагов от жука к оленю, но мы знаем, что по крайней мере одну последовательность можно построить, просто прослеживая предков жука в прошлое, к общему предку, и затем идя в восходящем порядке по аналогичной линии предков оленя.
Мы доказали, что существует линия пошаговых изменений, переходящая от жука к оленю, подразумевая наличие подобной линии от любого современного животного к любому другому такому же. Поэтому мы можем предполагать, что можно искусственно составить такой ряд давлений отбора, какой мог бы провести развитие по одной из таких линий. Этот быстрый мысленный эксперимент про линии позволил мне при обсуждении преобразований Д'Арси Томпсона говорить, что "В известном смысле очевидна строгая истинность того факта, что очертание любого животного можно преобразовать в очертания родственного животного посредством математического преобразования, хотя не очевидно, что преобразование будет простым". В этой книге я буду часто использовать технику мысленного эксперимента. Я предупреждаю читателя об этом заранее, так как учёных иногда раздражает недостаток реализма в таких формах рассуждений. Мысленные эксперименты и не обязаны быть реалистичными. Я полагаю, что они проясняют наши рассуждения о реальности.
Ещё одной особенностью жизни в этом мире, которую, подобно разделению на два пола, мы воспринимали как нечто само собой разумеющееся (хотя возможно зря), состоит в том, что живая материя группируется в дискретные пакеты, называемые организмами. В частности биологи, интересующиеся функциональными вопросами, обычно полагают соответствующей единицей обсуждения индивидуальный организм. Слово "конфликт", обычно означает конфликт между организмами, каждый из которых стремится максимизировать свою индивидуальную выживаемость. Мы признаём меньшие единицы - клетки и гены, и большие единицы - популяции, сообщества и экосистемы, но индивидуальный организм, - как дискретная единица, без сомнения мощно довлеет в умах зоологов, особенно интересующихся адаптивным значением поведения животных. Одна из целей этой книги состоит в разрушении эту довлеющей догмы. Я хочу убрать акцент с индивидуального тела как с центральной функциональной единицы обсуждения. По крайней мере, я хочу помочь вам осознать, насколько часто мы впадаем в иллюзию "само собой разумеющихся", вещей, когда мы смотрим на жизнь, как на набор дискретных индивидуальных организмов.
Поддерживаемый мною тезис состоит вот в чём. Законно говорить об адаптации как существующей "для выгоды" кое-чего, но это "кое-что" лучше не рассматривать как индивидуальный организм. Это - меньшая единица, которую я называю активным репликатором зародышевой линии. Наиболее важный вид репликатора - "ген" или маленький фрагмент генетического материала. Конечно, репликаторы подвергаются отбору не впрямую, а через "полномочных представителей" - в соответствии с оценкой их фенотипических эффектов. Хотя для некоторых целей удобно думать об этих фенотипических эффектах как о чём-то едином, спакетированном в дискретных "носителях", таких как индивидуальный организм, но в этом нет принципиальной необходимости. Скорее, о репликаторе следует думать, как о сущности, расширенной своими фенотипическими эффектами; весь мир в целом состоит из его эффектов, а не только тело особи, в котором ему случилось оказаться.
В поддержку можно привести пример мыслительного переключения, аналогичного кубу Неккера. Взглянув на жизнь, мы сначала видим совокупность взаимодействующих особей. Мы знаем, что они состоят из меньших единиц; мы также знаем, что они в свою очередь, есть части больших сложных единиц, но мы сосредотачиваем наш пристальный взгляд на особях как целом. Затем внезапно картина перещёлкивается. По-прежнему в ней есть особи; они никуда не делись, но они теперь кажутся прозрачными. Мы видим внутри их реплицирующиеся фрагменты ДНК; мы также видим окружающий эти особи мир, как арену, на которой эти генетические фрагменты продолжают состязания своих способностей воздействия. Гены управляют миром, и формируют его так, чтобы он помогал их репликации. Случилось так, что они "выбрали" основным путём изменения внешнего мира сборку материи в большие многоклеточные агрегаты, которыми мы называем организмами, но могло быть и иначе. Главное, что при этом происходит - копирование молекул, которые гарантируют своё выживание своим фенотипическим воздействием на мир. И лишь случайно получилось так, что эти фенотипические эффекты оказались спакетированы в модули, называемые индивидуальными организмами.
Сейчас мы не ценим организм за этот знаменательный феномен. О любом широко распространённом биологическом явлении мы привыкли спрашивать: "Какова его ценность для выживания?". Но мы не говорим, "Какова ценность для выживания жизни, упакованной в дискретные модули, называемые организмами?". Мы принимаем это как данность, присущую жизни. Как я уже отметил, организм автоматически становится предметом наших вопросов о ценности для выживания других сущностей: каким образом этот поведенческий паттерн приносит выгоду особи, выполняющей его? Каким образом эта морфологическая структура приносит пользу особи, её включающей?
Стало своего рода "центральной теоремой" (Barash 1977) современной этологии, что от особей следует ожидать такого поведения, которое приносит пользу их собственной итоговой приспособленности (Гамильтон io, 64a, b), но не пользу кому-то или чему-то другому. Мы не спрашиваем, каким образом поведение левой задней ноги приносит пользу левой задней ноге. Точно так же сейчас мало кто из нас спрашивает, как поведение группы особей, или экосистемы, выгодно этой группе или экосистеме. Мы рассматриваем группы и экосистемы как совокупности враждующих, или тревожно сожительствующих организмов; но мы рассматриваем ноги, почки, и клетки как сотрудничающие компоненты одного организма. Я не настаиваю на возражениях против фокусировки внимания на отдельных особях, но обращаю внимание на то, что мы считаем эту фокусировку само собой разумеющейся. Возможно нам нужно прекратить принимать это как очевидное и усомниться в особи, как в чём-то таком, что нуждается в объяснении "по его неотъемлемому праву", подобно нашим недавним обсуждениям полового размножения как чего-то такого, что нужно было объяснять также "по неотъемлемому праву".
В этом месте историческая неприятность биологии нуждается в утомительном отступлении от темы. Господствующая ортодоксия, описанная в предыдущем параграфе, центральная догма об особях, действующих в направлении максимизации своего репродуктивного успеха, парадигма "эгоистичного организма", выдвинутая Дарвином, продолжает доминировать и сегодня. Следовательно, кто-то может вообразить, что получил полное удовольствие за свои деньги и должен уже быть готовым к революциям, или по крайней мере достаточно окрепнуть, чтобы противостоять направленным против предрассудков булавочным уколам, вроде тех, что может наносить эта книга. К сожалению, и это как раз та историческая неприятность, о который я упомянул: хотя верно то, что желание обращать внимание на меньшие чем особь единицы, действующие во имя собственной выгоды возникает редко, но то же самое не всегда верно для единиц, бОльших особи. Годы, прошедшие со времён Дарвина, видели удивительное отступление от его сфокусированной на особи позиции - впадение в сентиментально-бессознательный групповой отбор, умело задокументированный Вильямсом (1966), Гизелином (1974a) и другими. Как выразился Гамильтон (1975a) "... Почти целый пласт биологии сломя голову ринулся в том направлении, куда Дарви вступил очень осторожно, или не вступал вообще". И только в последние годы, примерно совпавшие с запоздалым вхождением в моду собственных идей Гамильтона (Докинз 1979b), это сумасшедшее бегство было остановлено и перенаправлено. С тяжёлыми боями мы пробивались назад, изнурительно отстреливаясь от иезуитски искушённого и освящённого арьергарда новых групповых селекционистов, пока мы наконец не восстановили Дарвиновские позиции, которые я обозначаю меткой "эгоистичный организм"; позиции, которые в их современной форме доминируют в концепции "итоговой приспособленности". Есть тем не менее ощущение, от этой с трудом завоеванной твердыни я могу отказаться здесь - отказаться едва ли не ранее, чем она должным образом укреплена; и во имя чего? Во имя мерцающего куба Неккера, метафизической химеры под названием расширенный фенотип?
Нет, отказ от таких достижений далёк от моих намерений. Парадигма эгоистичного организма гораздо предпочтительнее того, что Гамильтон (1977) назвал "старой, умирающей парадигмой адаптации ради пользы вида". Толкование "расширенного фенотипа", как имеющего некую связь с адаптацией на уровне группы, будет ошибочным. Эгоистичный организм, и эгоистичный ген с его расширенным фенотипом - есть два представления одного и того же куба Неккера. Читатель не будет ощущать концептуального переключения, которому я стремлюсь потворствовать, если он изначально не видит правильный куб. Эта книга адресована тем, кто скорее одобряет ныне признанный взгляд на жизнь с эгоистичными организмами, чем взгляд какой-нибудь концепции "групповой выгоды".
Я не утверждаю, что концепция эгоистичного организма строго неправильна, но мой аргумент - в его резкой форме состоит в том, что она смотрит на мир неправильным способом. Однажды я подслушал разговор выдающегося Кембриджского этолога, с выдающимся Австрийским этологом (они спорили о развитии поведения): "Вы знаете, в самом-то деле мы соглашаемся. Вы лишь это неправильно говорите". О досточтимый "индивидуальный селекционист"! Мы действительно почти согласны, по крайней мере в споре с групповыми селекционистами. Вы лишь это неправильно видите!
Боннер (1958), рассуждая об одноклеточных организмах, говорил: "...Что такого особенного даёт этим организмам использование клеточно-ядерных генов? Как они возникали в ходе отбора?". Это - хороший пример образного, радикального вопроса, какой, думаю мы должны задавать о жизни вообще. Но если принять тезис этой книги, то конкретно этот вопрос нужно перевернуть вверх тормашками. Вместо выяснения того, что даёт организму использование клеточно-ядерных генов, мы должны спросить, почему гены захотели сгруппироваться в ядрах, и далее в организмах. В первых строках этой работы Боннер говорит: "Я не предполагаю высказывать что-то новое или оригинальное в этих лекциях. Но я - большой сторонник рассказывать о знакомых, известных вещах задом наперёд и кверху дном, в надежде глубже увидеть с этой точки обзора некоторые старые факты. Это похоже на рисование абстрактной картины вверх ногами - я не утверждаю, что смысл картины внезапно прояснится, но что-то ранее скрытое в структуре композиции может проявиться. Я натолкнулся на это высказывание, уже написав раздел про куб Неккера, и был восхищён, найдя те же самые взгляды, выраженные столь уважаемым автором.
Проблема с моим кубом Неккера, как и с абстрактной живописью Боннера, состоит в том, что как аналогии, они могут быть слишком робки и неамбициозны. Аналогия с кубом Неккера выражает мой минимум надежд на эту книгу. Я вполне уверен, что взгляд на жизнь в свете генетических репликаторов, выживающих с помощью их расширенных фенотипов, как минимум столь же удовлетворителен, сколь и взгляд на неё в свете эгоистичных организмов, максимизирующих свою итоговую приспособленность. Во многих случаях эти два способа рассмотрения жизни будут в сущности эквивалентны. Позже я покажу, что определение "итоговой приспособленности" сформулировано так, чтобы сделать по возможности эквивалентными выражения "особь максимизирует свою итоговую приспособленность" и "генетические репликаторы максимизируют свою выживаемость". Поэтому биолог должен стараться опробовать оба стиля мышления, и выбирать тот, который он или она предпочитает. Но это, как я сказал, только минимум надежд. Я буду обсуждать явления, к примеру, "мейотический драйв", объяснение которых ясно начертано на втором образе куба, но которые вообще не имеют смысла, если мы твёрдо удержим наш мысленный взгляд на другом образе - образе эгоистичного организма. Переход от моей робкой надежды в самые смелые мечты произойдёт, когда целые области биологии - изучение коммуникации животных, их построек и орудийной деятельности, паразитизма и симбиоза, экологии сообществ, - все-все взаимодействия между организмами и внутри их, будут в конечном счёте освещены новым светом в соответствии с доктриной расширенного фенотипа. Также как адвокат, я буду стараться делать упор на самом сильном из возможных случаев, что скорее означает повод для более смелых надежд, чем осторожных минимальных ожиданий.
Если в конечном счёте осознать эти грандиозные надежды, то возможно будут простительны и менее скромные аналогии, чем куб Неккера. Колин Турнбалл (1961) как-то вывел своего друга-пигмея Кенга впервые в его жизни из леса, они поднялись вместе на гору и взглянули на равнины. Кенг увидел несколько буйволов "лениво пасущихся в нескольких милях отсюда, далеко внизу. Он повернулся ко мне и спросил: "Что это там за букашки?"... Я не сразу понял, но потом догадался, что в лесу его был обзор столь ограничен, что ему не было особой потребности делать автоматическую поправку на расстояние при оценке размеров. Здесь, на равнине, Кенг по-видимому впервые видел бесконечные мили незнакомых полей, где не было деревьев, могущих дать ему базу для сравнения... Когда я сказал Кенгу, что эти букашки - буйволы, он взорвался от смеха и попросил меня не говорить такой глупой лжи..." (стр. 227-228).
В целом эта книга - работа адвоката, но плохого адвоката - такого, который опрометчиво перескакивает к своему заключению, когда присяжные ещё полны скепсиса. Второй образ моего куба Неккера не будет в чётком фокусе почти до окончания книги. Начальные главы готовят почву, стараясь предвосхитить риск определённых недоразумений, различными способами препарируют первый образ куба Неккера, показывая причины того, почему парадигма эгоистичной особи, пусть не полностью неправильна, но может привести к трудностям.
Части некоторых начальных глав - откровенная ретроспектива и даже оборона. Реакция на предыдущую работу (Докинз 1976a) наводит на мысль, что эта книга, вероятно, возбудит ненужное опасение за то, что пропагандирует два непопулярных "изма" - "генетический детерминизм" и "адаптационизм". Я и сам признаюсь, что меня раздражает книга, каждая страница которой вызывает у меня ворчание: "Да, но...", хотя автор мог бы легко предотвратить моё беспокойство небольшим тактичным объяснением вначале. Главы 2 и 3 стараются заблаговременно исключить по крайней мере два главных источника "да-нокания".
Глава 4 возбуждает судебное дело против эгоистичного организма, и начинает намекать на второй образ куба Неккера. Глава 5 возбуждает судебное дело в пользу "репликатора" как фундаментальной единицы естественного отбора. Глава 6 возвращается к особи, и показывает что ни она, ни что-то другое не может быть главным кандидатом на роль репликатора кроме маленького генетического фрагмента, квалифицируемого как истинный репликатор. Скорее, об особи нужно думать как о "носителе" репликаторов. Глава 7 отклоняется от методологии исследования. Глава 8 обращает внимание на некоторые труднопреодолимые аномалии концепции эгоистичного организма, а глава 9 продолжает тему. Глава 10 обсуждает различные варианты "персональной приспособленности", и заключает, что они сбивают с толку, и вероятно необязательны.
Главы 11, 12 и 13 - сердцевина книги. Они шаг за шагом развивают идею именно расширенного фенотипа, второй образ куба Неккера. Наконец, в главе 14, мы оглядываемся назад, со свежим любопытством смотрим на особь и задаёмся вопросом почему, в конце концов, она - такой важный уровень в иерархии жизни.